

Comparative expression profiling of Leishmania: modulation in gene expression between species and in different host genetic backgrounds
- PMID: 19582145
- PMCID: PMC2701600
- DOI: 10.1371/journal.pntd.0000476
Comparative expression profiling of Leishmania: modulation in gene expression between species and in different host genetic backgrounds
Abstract
Background: Genome sequencing of Leishmania species that give rise to a range of disease phenotypes in the host has revealed highly conserved gene content and synteny across the genus. Only a small number of genes are differentially distributed between the three species sequenced to date, L. major, L. infantum and L. braziliensis. It is not yet known how many of these genes are expressed in the disease-promoting intracellular amastigotes of these species or whether genes conserved between the species are differentially expressed in the host.
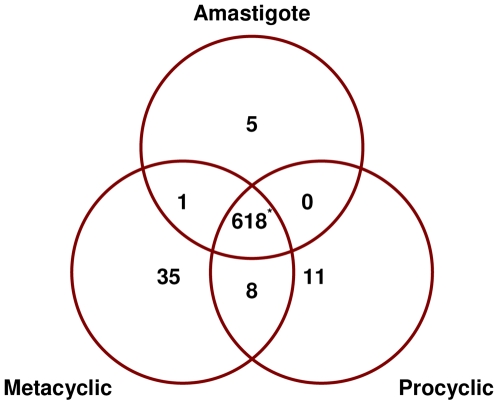
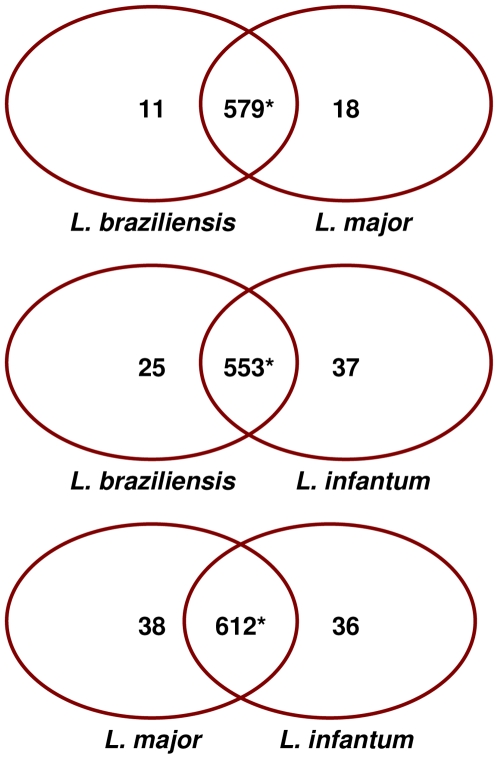
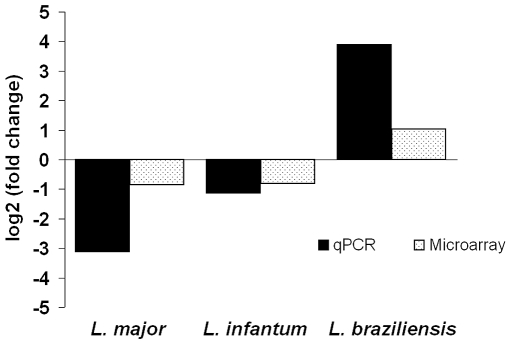
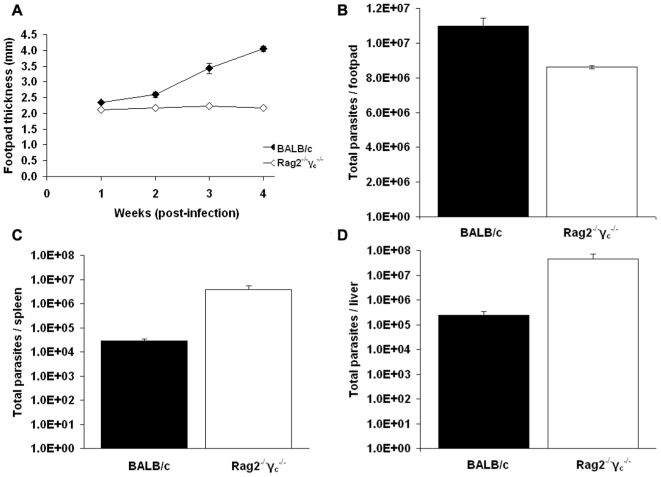
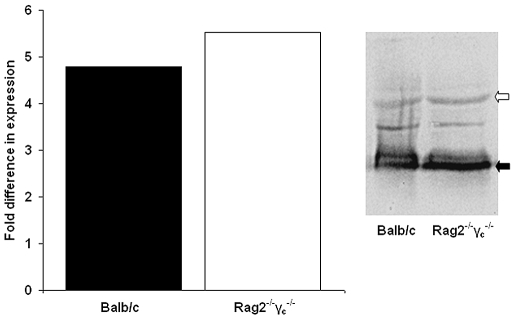
Methods/principal findings: We have used customised oligonucleotide microarrays to confirm that all of the differentially distributed genes identified by genome comparisons are expressed in intracellular amastigotes, with only a few of these subject to regulation at the RNA level. In the first large-scale study of gene expression in L. braziliensis, we show that only approximately 9% of the genes analysed are regulated in their RNA expression during the L. braziliensis life cycle, a figure consistent with that observed in other Leishmania species. Comparing amastigote gene expression profiles between species confirms the proposal that Leishmania transcriptomes undergo little regulation but also identifies conserved genes that are regulated differently between species in the host. We have also investigated whether host immune competence influences parasite gene expression, by comparing RNA expression profiles in L. major amastigotes derived from either wild-type (BALB/c) or immunologically compromised (Rag2(-/-) gamma(c) (-/-)) mice. While parasite dissemination from the site of infection is enhanced in the Rag2(-/-) gamma(c) (-/-) genetic background, parasite RNA expression profiles are unperturbed.
Conclusion/significance: These findings support the hypothesis that Leishmania amastigotes are pre-adapted for intracellular survival and undergo little dynamic modulation of gene expression at the RNA level. Species-specific parasite factors contributing to virulence and pathogenicity in the host may be limited to the products of a small number of differentially distributed genes or the differential regulation of conserved genes, either of which are subject to translational and/or post-translational controls.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Murray HW, Berman JD, Davies CR, Saravia NG. Advances in leishmaniasis. Lancet. 2005;366:1561–1577. - PubMed
-
- Mauricio IL, Stothard JR, Miles MA. The strange case of Leishmania chagasi. Parasitol Today. 2000;16:188–189. - PubMed
-
- Guerbouj S, Guizani I, Speybroeck N, Le Ray D, Dujardin JC. Genomic polymorphism of Leishmania infantum: a relationship with clinical pleomorphism? Infect Genet Evol. 2001;1:49–59. - PubMed
-
- McMahon-Pratt D, Alexander J. Does the Leishmania major paradigm of pathogenesis and protection hold for New World cutaneous leishmaniases or the visceral disease? Immunol Rev. 2004;201:206–224. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
