

PIKfyve regulation of endosome-linked pathways
- PMID: 19582903
- PMCID: PMC2723830
- DOI: 10.1111/j.1600-0854.2009.00915.x
PIKfyve regulation of endosome-linked pathways
Abstract
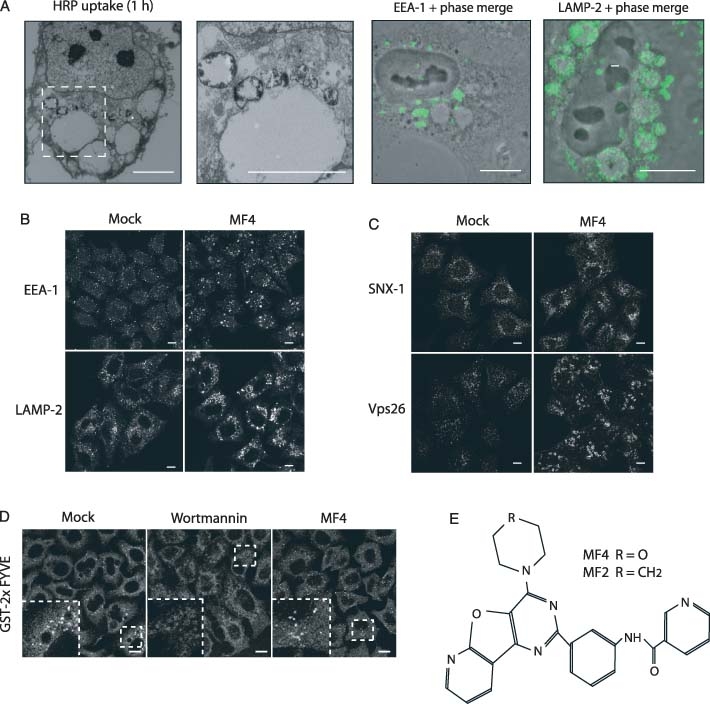
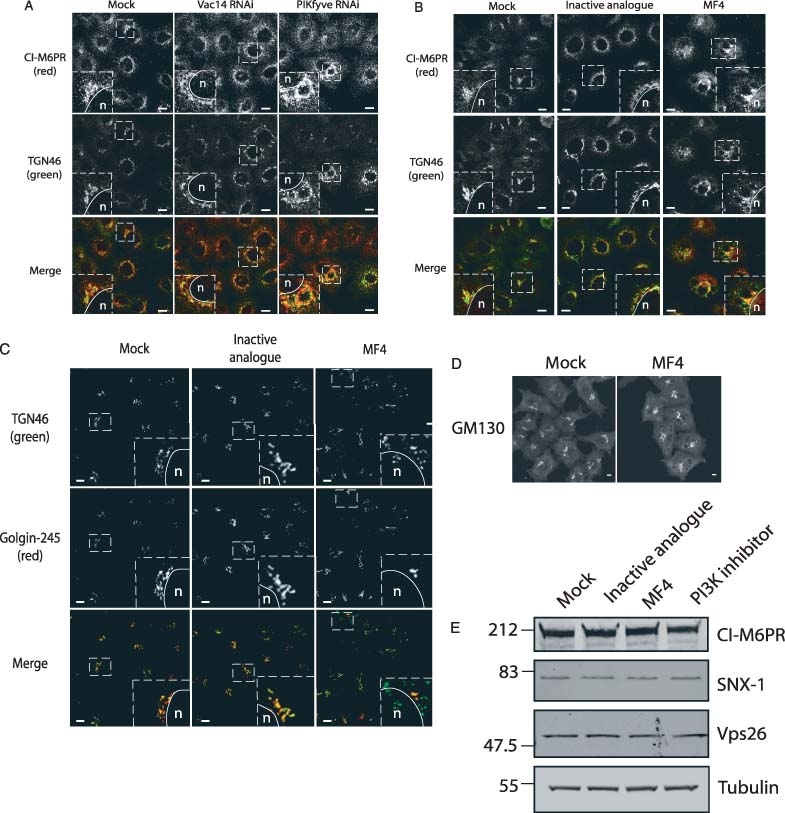
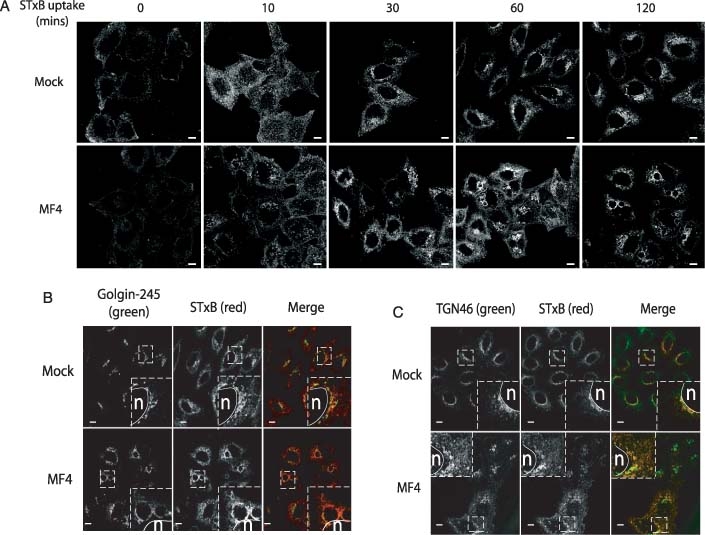
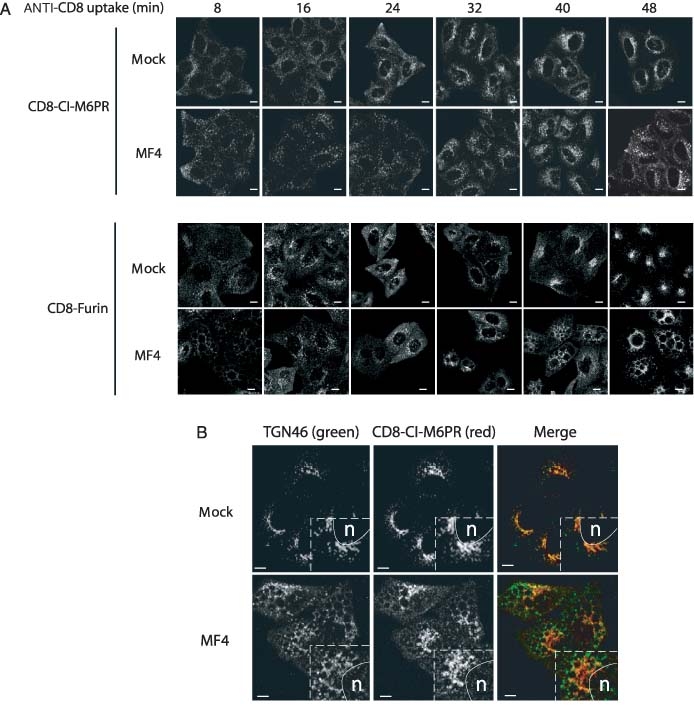
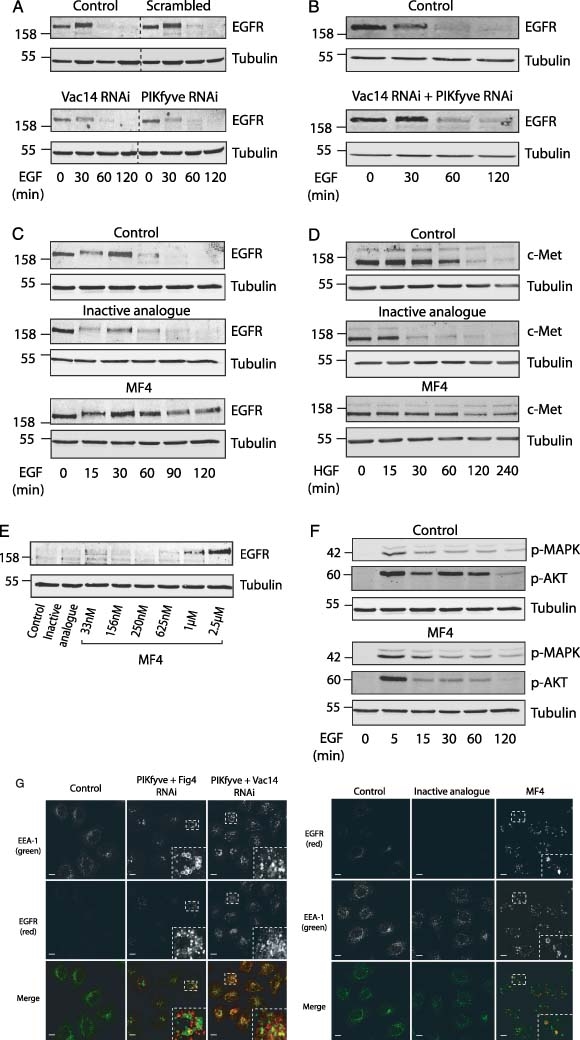
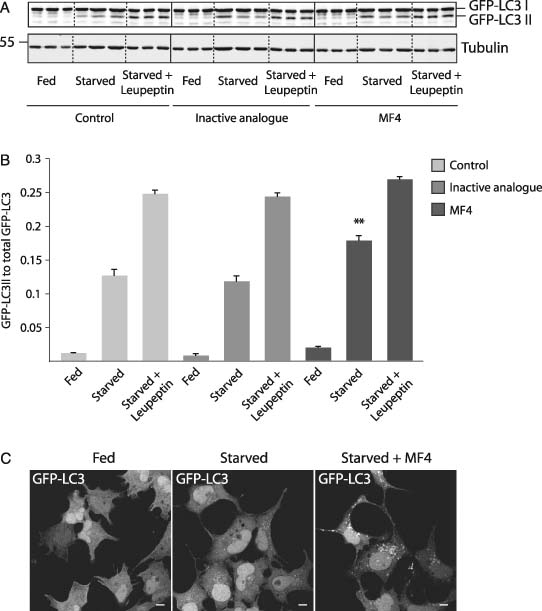
The phosphoinositide 5-kinase (PIKfyve) is a critical enzyme for the synthesis of PtdIns(3,5)P2, that has been implicated in various trafficking events associated with the endocytic pathway. We have now directly compared the effects of siRNA-mediated knockdown of PIKfyve in HeLa cells with a specific pharmacological inhibitor of enzyme activity. Both approaches induce changes in the distribution of CI-M6PR and trans-Golgi network (TGN)-46 proteins, which cycles between endosomes and TGN, leading to their accumulation in dispersed punctae, whilst the TGN marker golgin-245 retains a perinuclear disposition. Trafficking of CD8-CI-M6PR (retromer-dependent) and CD8-Furin (retromer-independent) chimeras from the cell surface to the TGN is delayed following drug administration, as is the transport of the Shiga toxin B-subunit. siRNA knockdown of PIKfyve produced no defect in epidermal growth factor receptor (EGFR) degradation, unless combined with knockdown of its activator molecule Vac14, suggesting that a low threshold of PtdIns(3,5)P2 is necessary and sufficient for this pathway. Accordingly pharmacological inhibition of PIKfyve results in a profound block to the lysosomal degradation of activated epidermal growth factor (EGF) and Met receptors. Immunofluorescence revealed EGF receptors to be trapped in the interior of a swollen endosomal compartment. In cells starved of amino acids, PIKfyve inhibition leads to the accumulation of the lipidated form of GFP-LC3, a marker of autophagosomal structures, which can be visualized as fluorescent punctae. We suggest that PIKfyve inhibition may render the late endosome/lysosome compartment refractory to fusion with both autophagosomes and with EGFR-containing multivesicular bodies.
Figures
References
-
- Lindmo K, Stenmark H. Regulation of membrane traffic by phosphoinositide 3-kinases. J Cell Sci. 2006;119:605–614. - PubMed
-
- Joly M, Kazlauskas A, Corvera S. Phosphatidylinositol 3-kinase activity is required at a postendocytic step in platelet-derived growth-factor receptor trafficking. J Biol Chem. 1995;270:13225–13230. - PubMed
-
- Mills IG, Jones AT, Clague MJ. Involvement of the endosomal autoantigen EEA1 in homotypic fusion of early endosomes. Curr Biol. 1998;8:881–884. - PubMed
-
- Raiborg C, Bache KG, Gillooly DJ, Madshus IH, Stang E, Stenmark H. Hrs sorts ubiquitinated proteins into clathrin-coated microdomains of early endosomes. Nat Cell Biol. 2002;4:394–398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
