

Cell biology and physiology of the uroepithelium
- PMID: 19587142
- PMCID: PMC2801337
- DOI: 10.1152/ajprenal.00327.2009
Cell biology and physiology of the uroepithelium
Abstract
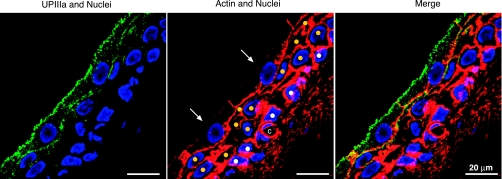
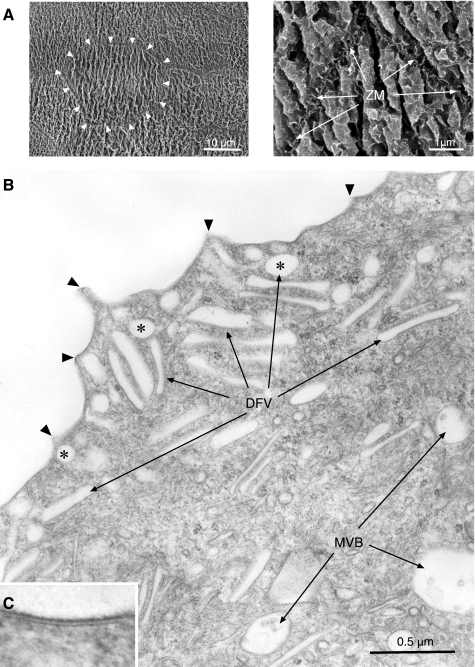
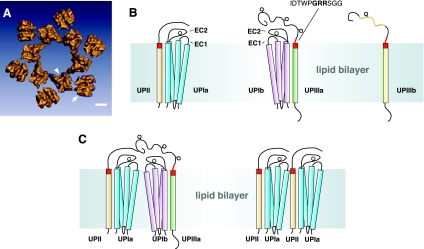
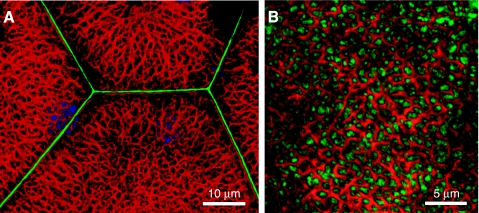
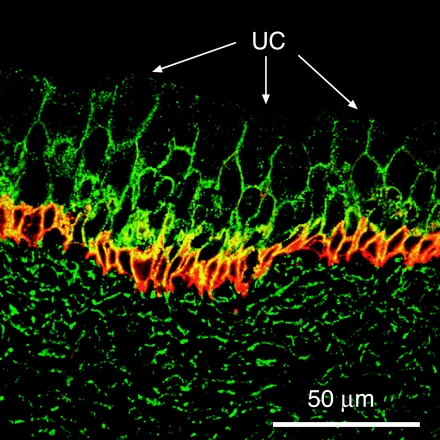
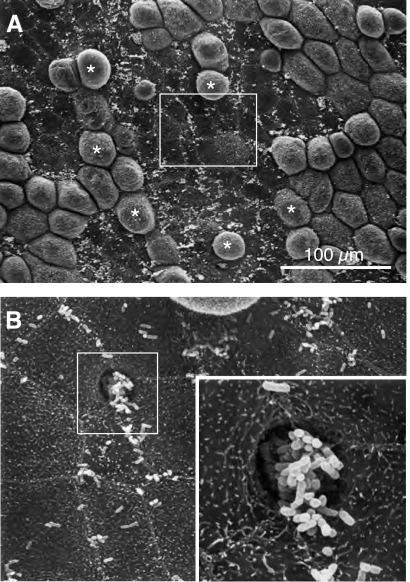
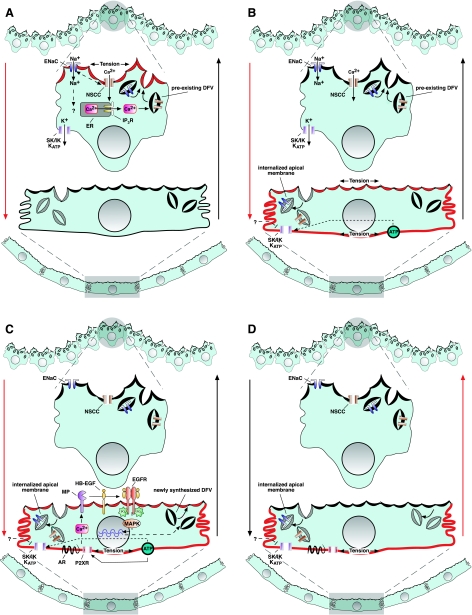
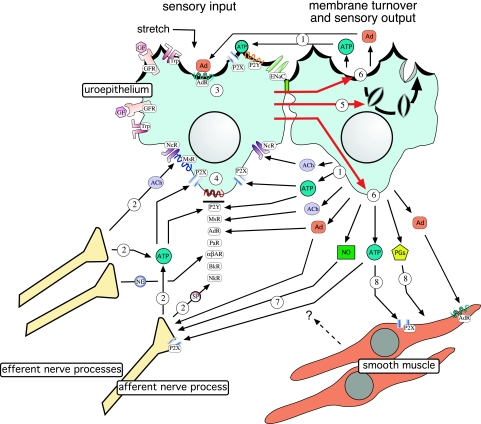
The uroepithelium sits at the interface between the urinary space and underlying tissues, where it forms a high-resistance barrier to ion, solute, and water flux, as well as pathogens. However, the uroepithelium is not simply a passive barrier; it can modulate the composition of the urine, and it functions as an integral part of a sensory web in which it receives, amplifies, and transmits information about its external milieu to the underlying nervous and muscular systems. This review examines our understanding of uroepithelial regeneration and how specializations of the outermost umbrella cell layer, including tight junctions, surface uroplakins, and dynamic apical membrane exocytosis/endocytosis, contribute to barrier function and how they are co-opted by uropathogenic bacteria to infect the uroepithelium. Furthermore, we discuss the presence and possible functions of aquaporins, urea transporters, and multiple ion channels in the uroepithelium. Finally, we describe potential mechanisms by which the uroepithelium can transmit information about the urinary space to the other tissues in the bladder proper.
Figures
References
-
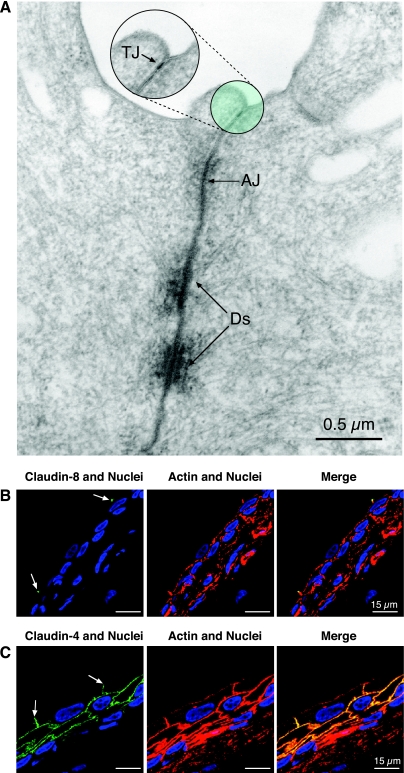
- Acharya P, Beckel J, Wang E, Ruiz W, Rojas R, Apodaca G. Distribution of the tight junction proteins ZO-1, occludin, and claudin-4, -8, and -12 in bladder epithelium. Am J Physiol Renal Physiol 287: F305–F318, 2004 - PubMed
-
- Adachi W, Okubo K, Kinoshita S. Human uroplakin Ib in ocular surface epithelium. Invest Ophthalmol Vis Sci 41: 2900–2905, 2000 - PubMed
-
- Alenghat FJ, Nauli SM, Kolb R, Zhou J, Ingber DE. Global cytoskeletal control of mechanotransduction in kidney epithelial cells. Exp Cell Res 301: 23–30, 2004 - PubMed
-
- Alroy J, Merk FB, Morre DJ, Weinstein RS. Membrane differentiation in the Golgi apparatus of mammalian urinary bladder epithelium. Anat Rec 203: 429–440, 1982 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources