

Deletion of the alpha 7 nicotinic acetylcholine receptor gene improves cognitive deficits and synaptic pathology in a mouse model of Alzheimer's disease
- PMID: 19587288
- PMCID: PMC2753494
- DOI: 10.1523/JNEUROSCI.6159-08.2009
Deletion of the alpha 7 nicotinic acetylcholine receptor gene improves cognitive deficits and synaptic pathology in a mouse model of Alzheimer's disease
Abstract
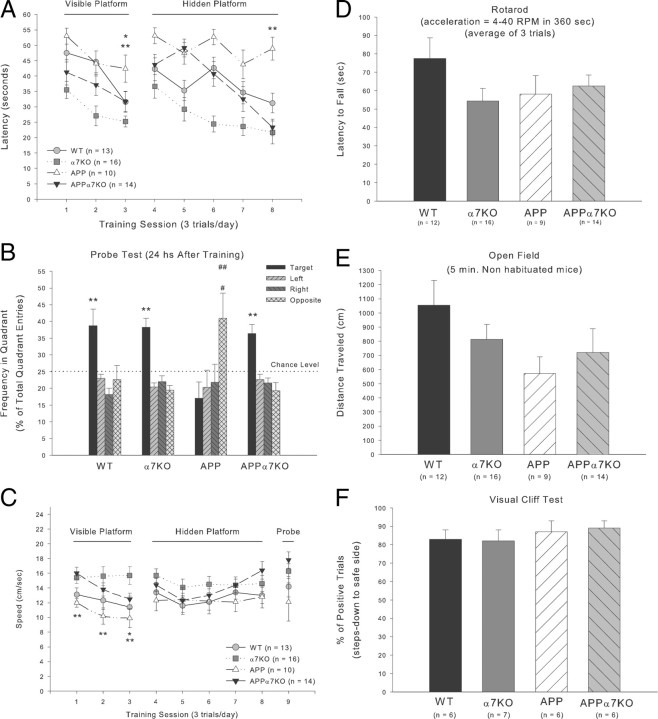
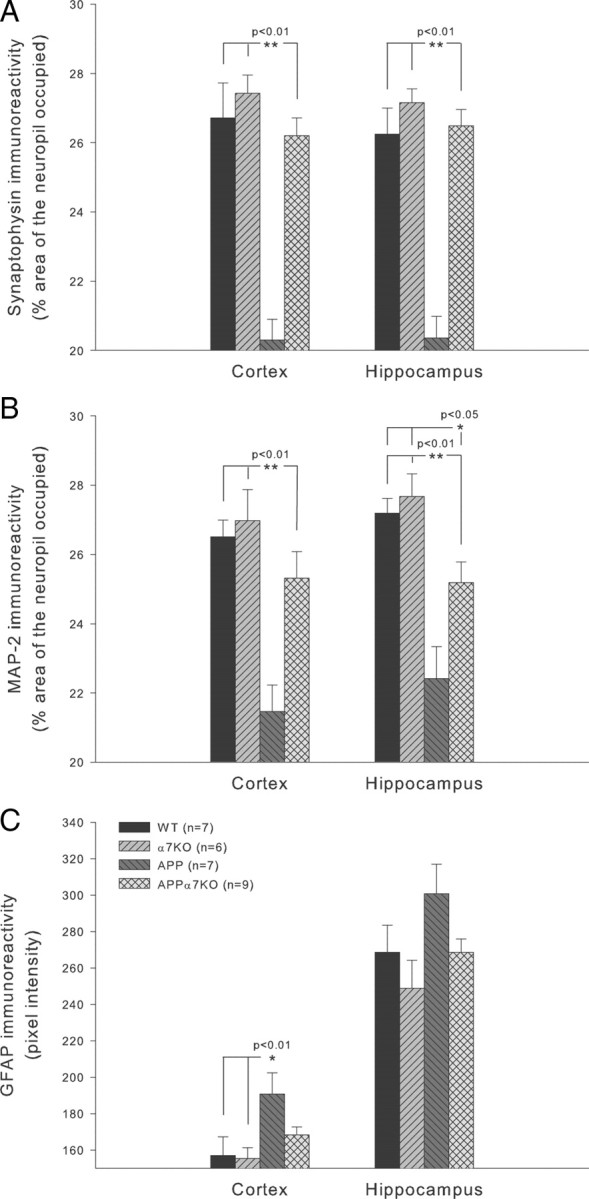
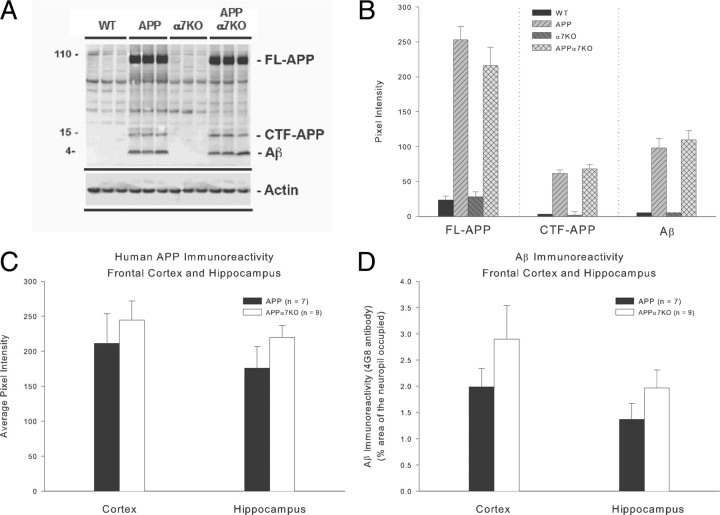
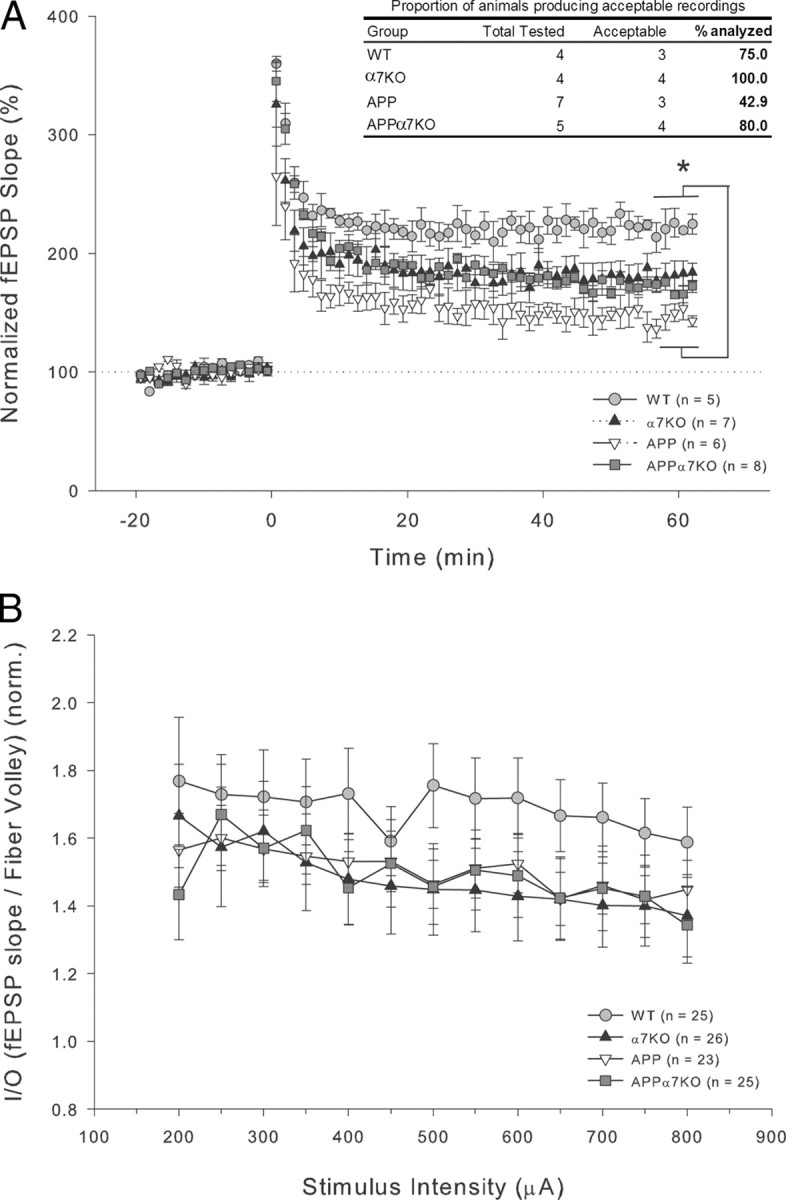
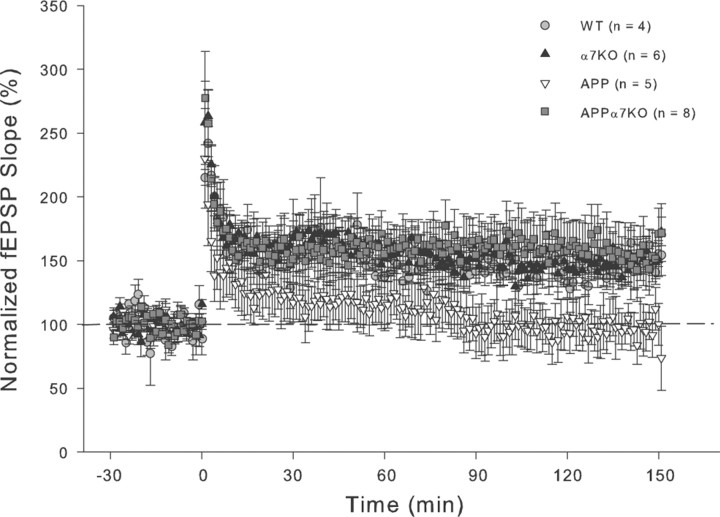
It has been recently shown that the Alzheimer's disease (AD) pathogenic peptide amyloid beta(1-42) (Abeta(1-42)) binds to the alpha7 nicotinic acetylcholine receptor (alpha7nAChR) with high affinity and the alpha7nAChR and Abeta(1-42) are both found colocalized in neuritic plaques of human brains with AD. Moreover, the intraneuronal accumulation of Abeta(1-42) was shown to be facilitated by its high-affinity binding to the alpha7nAChR, and alpha7nAChR activation mediates Abeta-induced tau protein phosphorylation. To test the hypothesis that alpha7nAChRs are involved in AD pathogenesis, we used a transgenic mouse model of AD overexpressing a mutated form of the human amyloid precursor protein (APP) and lacking the alpha7nAChR gene (APPalpha7KO). We have shown that, despite the presence of high amounts of APP and amyloid deposits, deleting the alpha7nAChR subunit in the mouse model of AD leads to a protection from the dysfunction in synaptic integrity (pathology and plasticity) and learning and memory behavior. Specifically, APPalpha7KO mice express APP and Abeta at levels similar to APP mice, and yet they were able to solve a cognitive challenge such as the Morris water maze test significantly better than APP, with performances comparable to control groups. Moreover, deleting the alpha7nAChR subunit protected the brain from loss of the synaptic markers synaptophysin and MAP2, reduced the gliosis, and preserved the capacity to elicit long-term potentiation otherwise deficient in APP mice. These results are consistent with the hypothesis that the alpha7nAChR plays a role in AD and suggest that interrupting alpha7nAChR function could be beneficial in the treatment of AD.
Figures
References
-
- Aracava Y, Pereira EF, Maelicke A, Albuquerque EX. Memantine blocks alpha7* nicotinic acetylcholine receptors more potently than N-methyl-d-aspartate receptors in rat hippocampal neurons. J Pharmacol Exp Ther. 2005;312:1195–1205. - PubMed
-
- Banerjee P, Samoriski G, Gupta S. Comments on “Memantine blocks {alpha}7* nicotinic acetylcholine receptors more potently than N-methyl-d-aspartate receptors in rat hippocampal neurons.”. J Pharmacol Exp Ther. 2005;313:928–929. author reply 930–933. - PubMed
-
- Bednar I, Paterson D, Marutle A, Pham TM, Svedberg M, Hellström-Lindahl E, Mousavi M, Court J, Morris C, Perry E, Mohammed A, Zhang X, Nordberg A. Selective nicotinic receptor consequences in APP(SWE) transgenic mice. Mol Cell Neurosci. 2002;20:354–365. - PubMed
-
- Billings LM, Oddo S, Green KN, McGaugh JL, LaFerla FM. Intraneuronal Abeta causes the onset of early Alzheimer's disease-related cognitive deficits in transgenic mice. Neuron. 2005;45:675–688. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases