

Endothelial cell protein C receptor cellular localization and trafficking: potential functional implications
- PMID: 19587380
- PMCID: PMC2738580
- DOI: 10.1182/blood-2009-03-208900
Endothelial cell protein C receptor cellular localization and trafficking: potential functional implications
Abstract
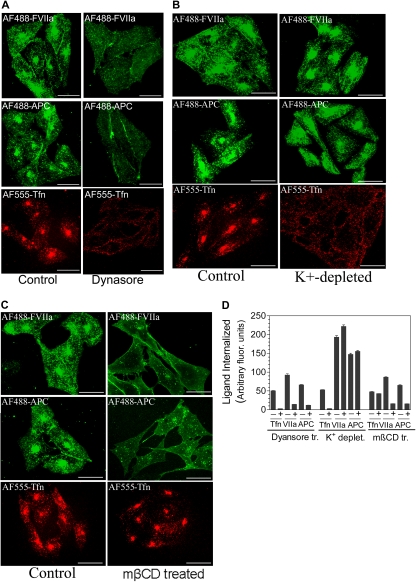
Although the binding of endothelial cell protein C receptor (EPCR) to its ligands is well characterized at the biochemical level, it remains unclear how EPCR interaction with its ligands at the cell surface impacts its cellular trafficking. We characterized the cellular localization and trafficking of EPCR in endothelial cells and a heterologous expression system. Immunofluorescence confocal microscopy studies revealed that a majority of EPCR is localized on the cell surface in membrane microdomains that are positive for caveolin-1. A small fraction of EPCR is also localized intracellularly in the recycling compartment. Factor VIIa (FVIIa) or activated protein C binding to EPCR promoted the internalization of EPCR. EPCR and EPCR-bound ligands were endocytosed rapidly via a dynamin- and caveolar-dependent pathway. The endocytosed receptor-ligand complexes were accumulated in a recycling compartment before being targeted back to the cell surface. EPCR-mediated FVIIa endocytosis/recycling also resulted in transport of FVIIa from the apical to the basal side. In vivo studies in mice showed that blockade of EPCR with EPCR-blocking antibodies impaired the early phase of FVIIa clearance. Overall, our results show that FVIIa or activated protein C binding to EPCR promotes EPCR endocytosis, and EPCR-mediated endocytosis may facilitate the transcytosis of FVIIa and its clearance from the circulation.
Figures







 ], adventitia and extravascular tissue.
], adventitia and extravascular tissue.References
-
- Regan LM, Stearns-Kurosawa DJ, Kurosawa S, et al. The endothelial cell protein C receptor: inhibition of activated protein C anticoagulant function without modulation of reaction with proteinase inhibitors. J Biol Chem. 1996;271(29):17499–17503. - PubMed
-
- Laszik Z, Mitro A, Taylor FB, Jr, Ferrell G, Esmon CT. Human protein C receptor is present primarily on endothelium of large blood vessels: implications for the control of the protein C pathway. Circulation. 1997;96(10):3633–3640. - PubMed
-
- Esmon CT. The endothelial protein C receptor. Curr Opin Hematol. 2006;13(5):382–385. - PubMed
-
- Riewald M, Petrovan RJ, Donner A, Mueller BM, Ruf W. Activation of endothelial cell protease activated receptor 1 by protein C pathway. Science. 2002;296(5574):1880–1882. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
