

The TLR9-MyD88 pathway is critical for adaptive immune responses to adeno-associated virus gene therapy vectors in mice
- PMID: 19587448
- PMCID: PMC2719948
- DOI: 10.1172/JCI37607
The TLR9-MyD88 pathway is critical for adaptive immune responses to adeno-associated virus gene therapy vectors in mice
Abstract
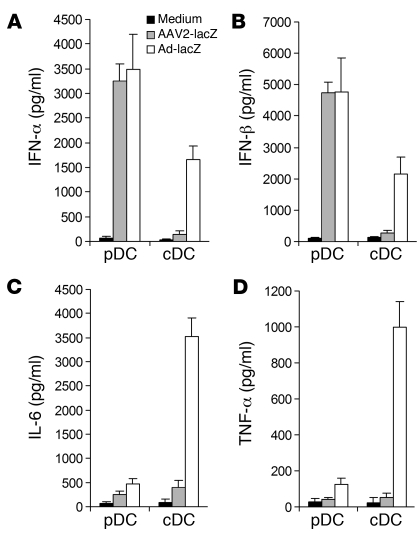
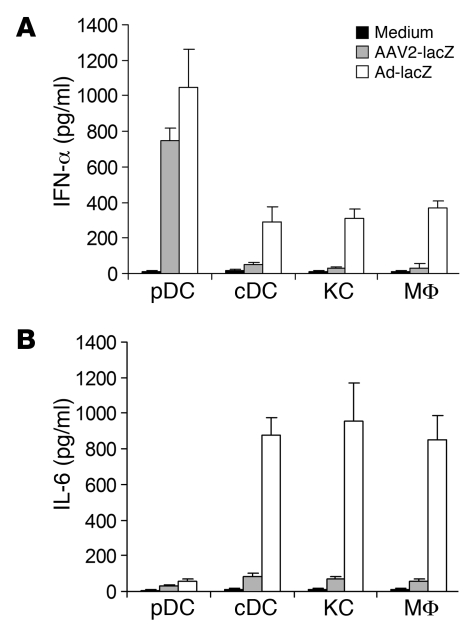
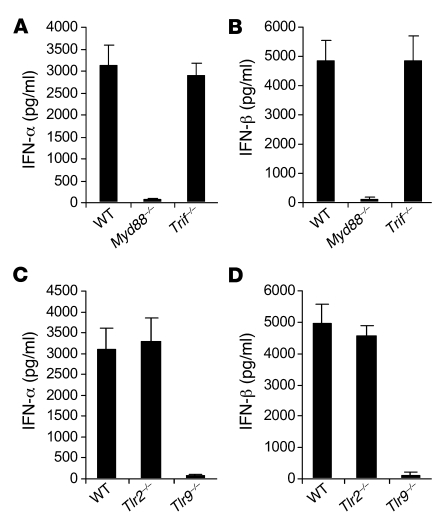
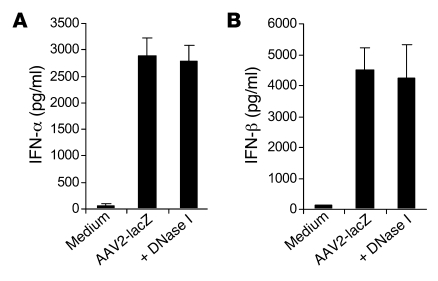
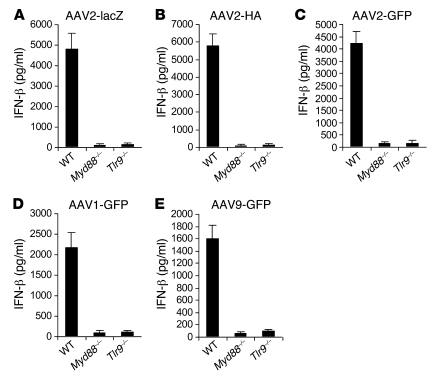
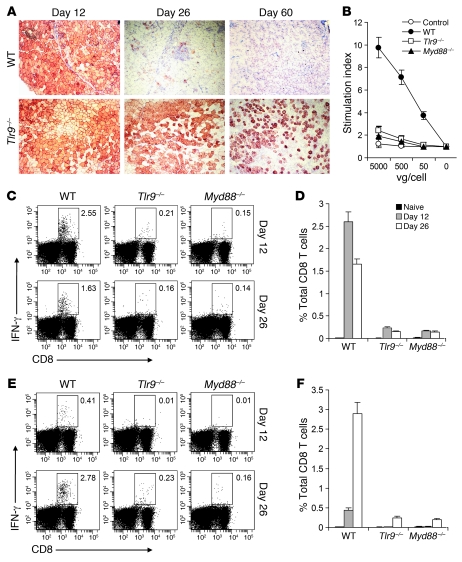
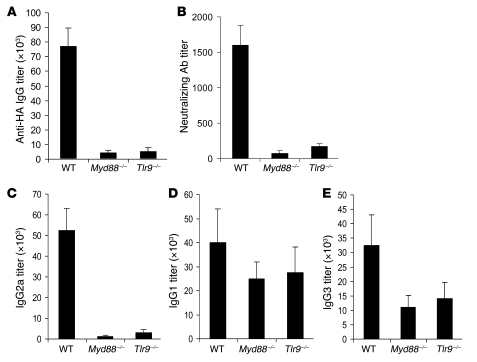
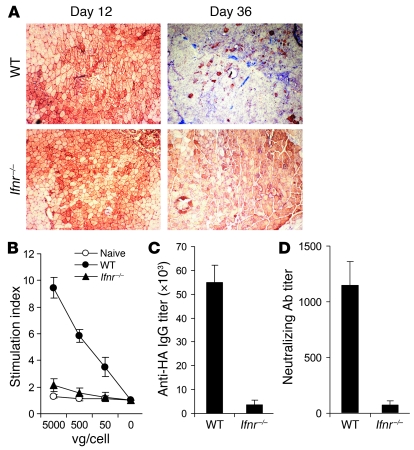
Recombinant adeno-associated viruses (AAVs) have been used widely for in vivo gene therapy. However, adaptive immune responses to AAV have posed a significant hurdle in clinical application of AAV vectors. Recent advances have suggested a crucial role for innate immunity in shaping adaptive immune responses. How AAV activates innate immunity, and thereby promotes AAV-targeted adaptive immune responses, remains unknown. Here we show that AAV activates mouse plasmacytoid DCs (pDCs) via TLR9 to produce type I IFNs. In vivo, the TLR9-MyD88 pathway was crucial to the activation of CD8+ T cell responses to both the transgene product and the AAV capsid, leading to loss of transgene expression and the generation of transgene product-specific and AAV-neutralizing antibodies. We further demonstrate that TLR9-dependent activation of adaptive immunity targeting AAV was mediated by type I IFNs and that human pDCs could be activated in vitro to induce type I IFN production via TLR9. These results reveal an essential role for the TLR9-MyD88-type I IFN pathway in induction of adaptive immune responses to AAV and suggest that strategies that interfere with this pathway may improve the outcome of AAV-mediated gene therapy in humans.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
