

Tissue elasticity and the ageing elastic fibre
- PMID: 19588272
- PMCID: PMC2813052
- DOI: 10.1007/s11357-009-9103-6
Tissue elasticity and the ageing elastic fibre
Abstract
The ability of elastic tissues to deform under physiological forces and to subsequently release stored energy to drive passive recoil is vital to the function of many dynamic tissues. Within vertebrates, elastic fibres allow arteries and lungs to expand and contract, thus controlling variations in blood pressure and returning the pulmonary system to a resting state. Elastic fibres are composite structures composed of a cross-linked elastin core and an outer layer of fibrillin microfibrils. These two components perform distinct roles; elastin stores energy and drives passive recoil, whilst fibrillin microfibrils direct elastogenesis, mediate cell signalling, maintain tissue homeostasis via TGFβ sequestration and potentially act to reinforce the elastic fibre. In many tissues reduced elasticity, as a result of compromised elastic fibre function, becomes increasingly prevalent with age and contributes significantly to the burden of human morbidity and mortality. This review considers how the unique molecular structure, tissue distribution and longevity of elastic fibres pre-disposes these abundant extracellular matrix structures to the accumulation of damage in ageing dermal, pulmonary and vascular tissues. As compromised elasticity is a common feature of ageing dynamic tissues, the development of strategies to prevent, limit or reverse this loss of function will play a key role in reducing age-related morbidity and mortality.
Figures
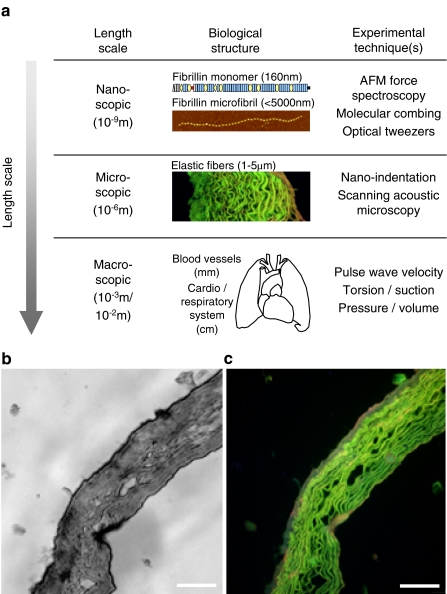
References
-
- Aaron BB, Gosline JM. Elastin as a random-network elastomer: a mechanical and optical analysis of single elastin fibers. Biopolymers. 1981;20:1247–1260.
-
- Agache PG, Monneur C, Leveque JL, Derigal J. Mechanical-properties and youngs modulus of human-skin in vivo. Arch Dermatol Res. 1980;269:221–232. - PubMed
-
- Andreotti L, Bussotti A, Cammelli D, Aiello E, Sampognaro S. Connective-tissue in aging lung. Gerontology. 1983;29:377–387. - PubMed
-
- Antoniou J, Steffen T, Nelson F, Winterbottom N, Hollander AP, Poole RA, Aebi M, Alini M. The human lumbar intervertebral disc - Evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J Clin Invest. 1996;98:996–1003. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials