

The flexibility of a distant loop modulates active site motion and product release in ribonuclease A
- PMID: 19588901
- PMCID: PMC2741010
- DOI: 10.1021/bi900830g
The flexibility of a distant loop modulates active site motion and product release in ribonuclease A
Abstract
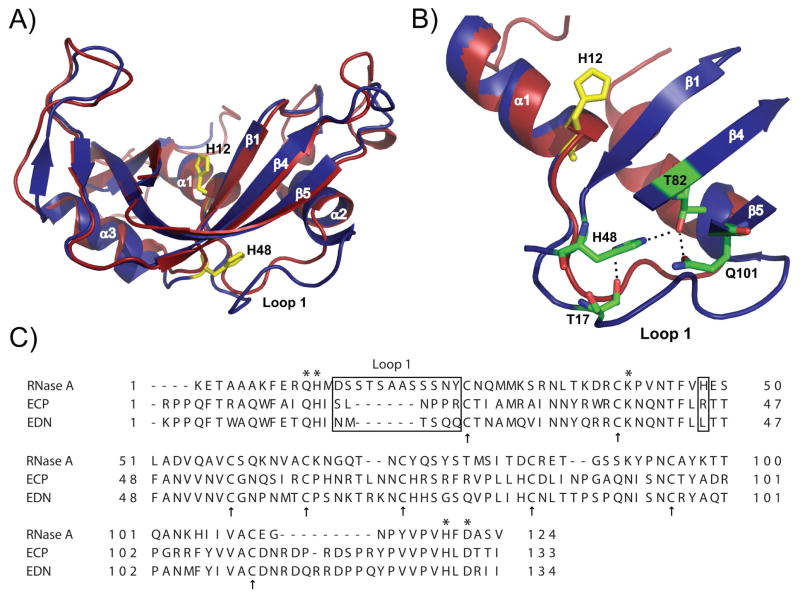
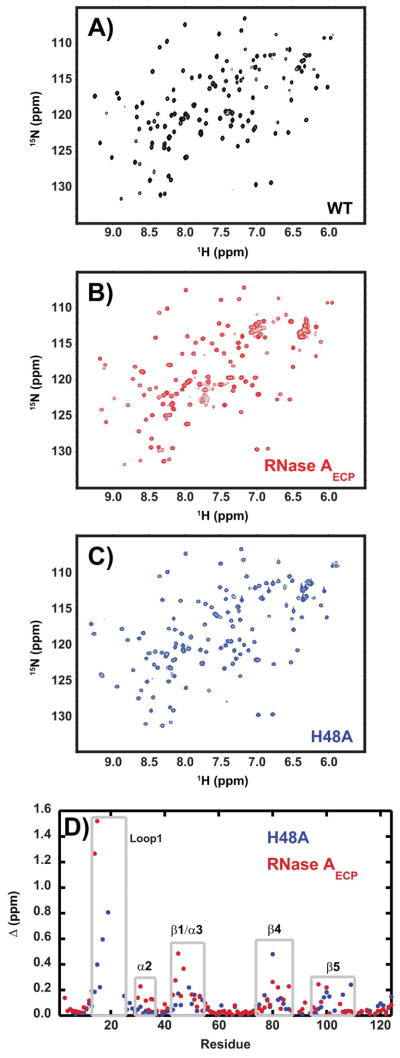
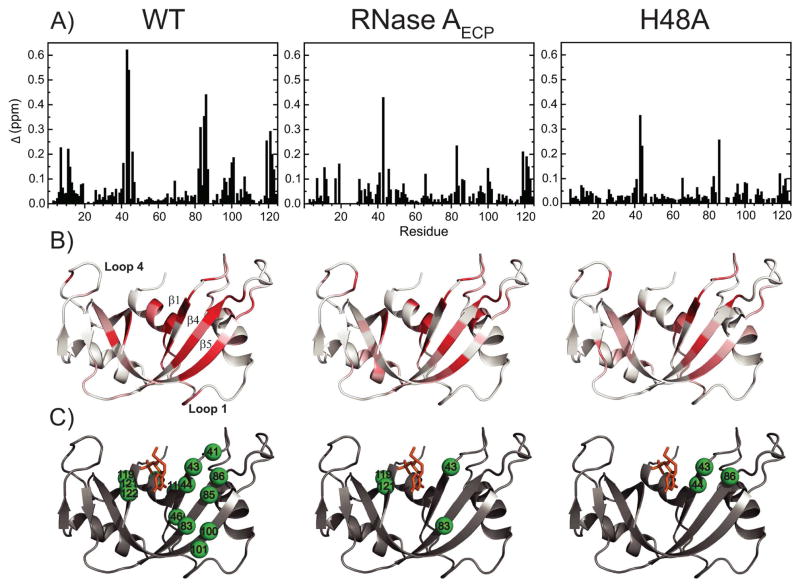
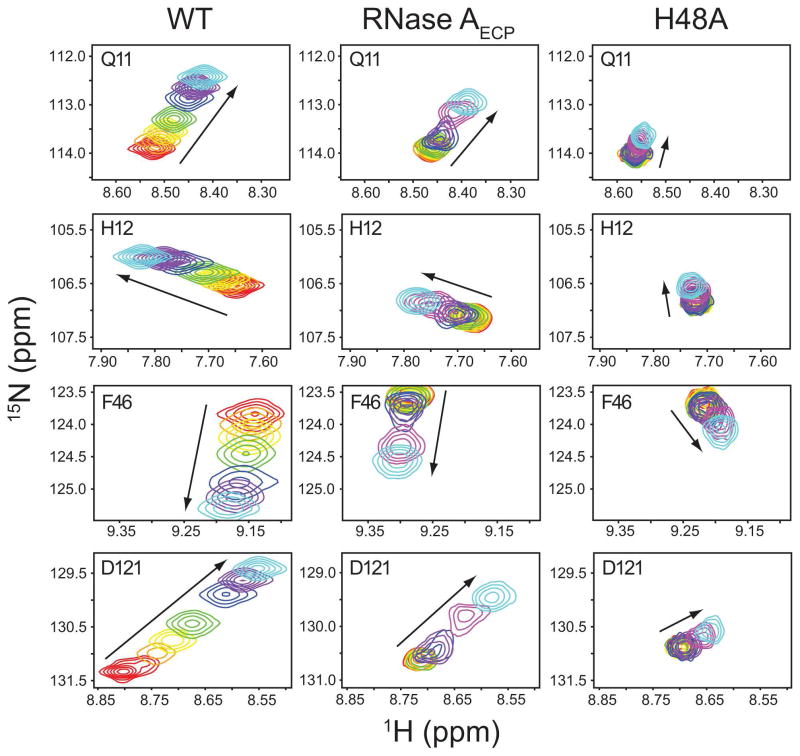
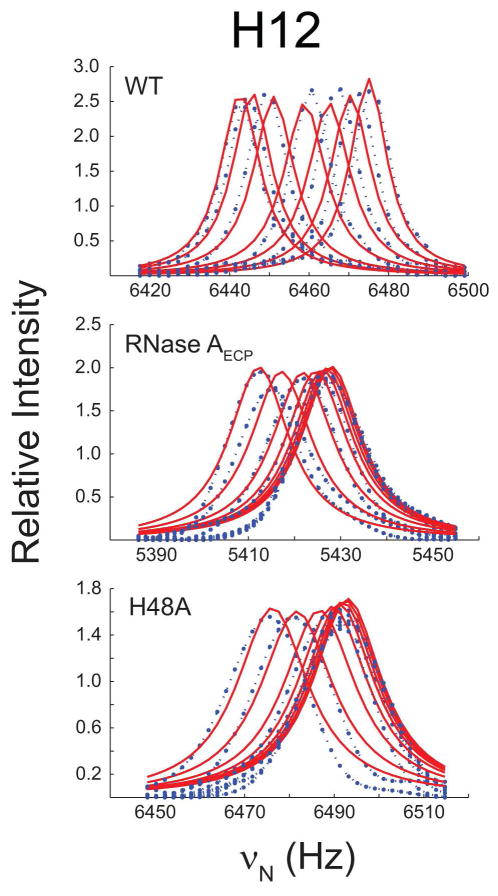
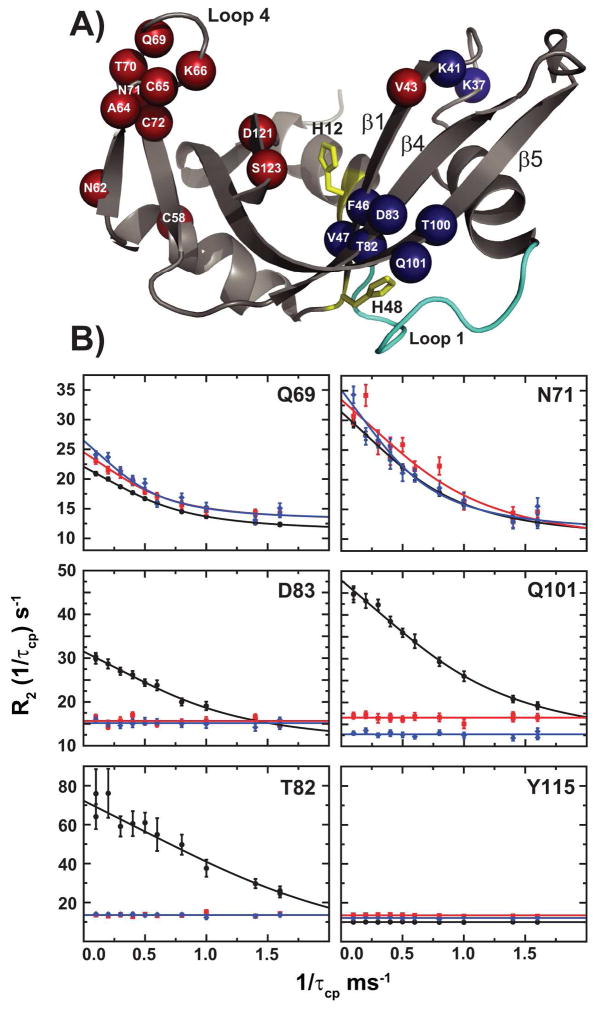
The role of the flexible loop 1 in protein conformational motion and in the dissociation of enzymatic product from ribonuclease A (RNase A) was investigated by creation of a chimeric enzyme in which a 6-residue loop 1 from the RNase A homologue, eosinophil cationic protein (ECP), replaced the 12-residue loop 1 in RNase A. The chimera (RNase A(ECP)) experiences only local perturbations in NMR backbone chemical shifts compared to WT RNase A. Many of the flexible residues that were previously identified in WT as involved in an important conformational change now experience no NMR-detected millisecond motions in the chimera. Likewise, binding of the product analogue, 3'-CMP, to RNase A(ECP) results in only minor chemical shift changes in the enzyme similar to what is observed for the H48A mutant of RNase A and in contrast to WT enzyme. For both RNase A(ECP) and H48A there is a 10-fold decrease in the product release rate constant, k(off), compared to WT, in agreement with previous studies indicating the importance of flexibility in RNase A in the overall rate-limiting product release step. Together, these NMR and biochemical experiments provide additional insight into the mechanism of millisecond motions in the RNase A catalytic cycle.
Figures
References
-
- Daniel RM, Dunn RV, Finney JL, Smith JC. The role of dynamics in enzyme activity. Annu Rev Biophys Biomol Struct. 2003;32:69–92. - PubMed
-
- Eisenmesser EZ, Millet O, Labeikovsky W, Korzhnev DM, Wolf-Watz M, Bosco DA, Skalicky JJ, Kay LE, Kern D. Intrinsic dynamics of an enzyme underlies catalysis. Nature. 2005;438:117–121. - PubMed
-
- Kovrigin EL, Loria JP. Enzyme dynamics along the reaction coordinate: critical role of a conserved residue. Biochemistry. 2006;45:2636–2647. - PubMed
-
- Boehr DD, McElheny D, Dyson HJ, Wright PE. The dynamic energy landscape of dihydrofolate reductase catalysis. Science. 2006;313:1638–1642. - PubMed
-
- Blacklow SC, Raines RT, Lim WA, Zamore PD, Knowles JR. Triosephosphate isomerase catalysis is diffusion controlled. Biochemistry. 1988;27:1158–1167. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
