

Epidithiodiketopiperazines block the interaction between hypoxia-inducible factor-1alpha (HIF-1alpha) and p300 by a zinc ejection mechanism
- PMID: 19589782
- PMCID: PMC2785371
- DOI: 10.1074/jbc.M109.009498
Epidithiodiketopiperazines block the interaction between hypoxia-inducible factor-1alpha (HIF-1alpha) and p300 by a zinc ejection mechanism
Abstract
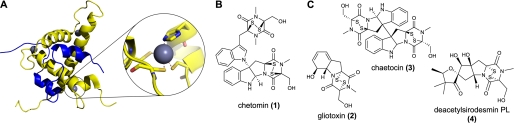
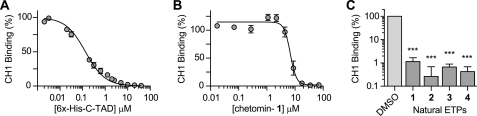
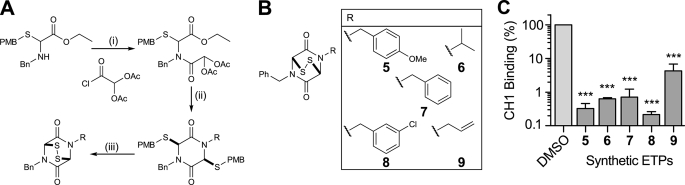
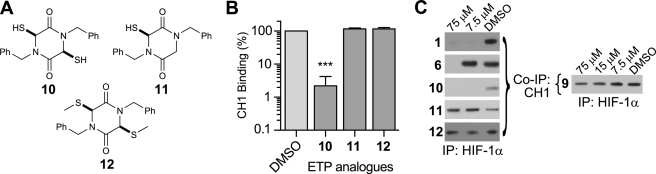
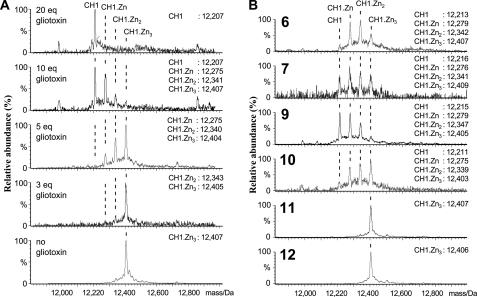
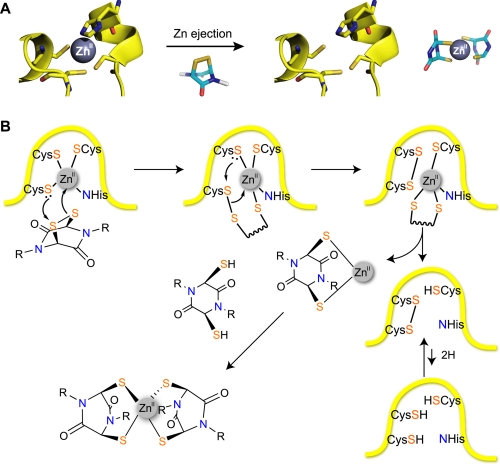
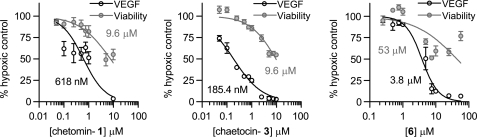
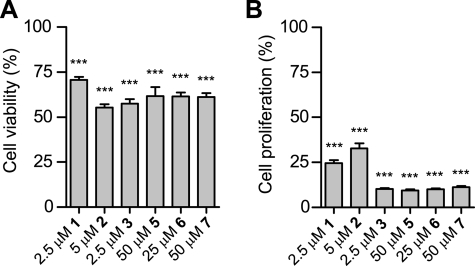
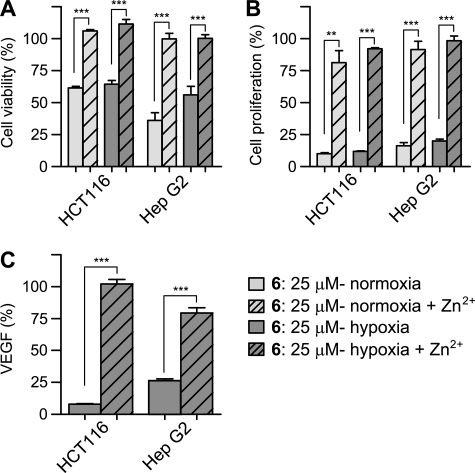
The hypoxic response in humans is regulated by the hypoxia-inducible transcription factor system; inhibition of hypoxia-inducible factor (HIF) activity has potential for the treatment of cancer. Chetomin, a member of the epidithiodiketopiperazine (ETP) family of natural products, inhibits the interaction between HIF-alpha and the transcriptional coactivator p300. Structure-activity studies employing both natural and synthetic ETP derivatives reveal that only the structurally unique ETP core is required and sufficient to block the interaction of HIF-1alpha and p300. In support of both cell-based and animal work showing that the cytotoxic effect of ETPs is reduced by the addition of Zn(2+) through an unknown mechanism, our mechanistic studies reveal that ETPs react with p300, causing zinc ion ejection. Cell studies with both natural and synthetic ETPs demonstrated a decrease in vascular endothelial growth factor and antiproliferative effects that were abrogated by zinc supplementation. The results have implications for the design of selective ETPs and for the interaction of ETPs with other zinc ion-binding protein targets involved in gene expression.
Figures
References
-
- Hirota K., Semenza G. L. (2006) Crit. Rev. Oncol. Hematol. 59, 15–26 - PubMed
-
- Kaelin W. G., Jr., Ratcliffe P. J. (2008) Mol. Cell 30, 393–402 - PubMed
-
- Lando D., Peet D. J., Whelan D. A., Gorman J. J., Whitelaw M. L. (2002) Science 295, 858–861 - PubMed
-
- Kung A. L., Wang S., Klco J. M., Kaelin W. G., Livingston D. M. (2000) Nat. Med. 6, 1335–1340 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
