

Expression of latent human immunodeficiency type 1 is induced by novel and selective histone deacetylase inhibitors
- PMID: 19590405
- PMCID: PMC3809117
- DOI: 10.1097/QAD.0b013e32832ec1dc
Expression of latent human immunodeficiency type 1 is induced by novel and selective histone deacetylase inhibitors
Abstract
Objectives: A family of histone deacetylases (HDACs) mediates chromatin remodeling, and repression of gene expression. Deacetylation of histones within the HIV-1 long terminal repeat (LTR) by HDACs plays a key role in the maintenance of latency, whereas acetylation of histones about the LTR is linked to proviral expression and escape of HIV from latency. Global HDAC inhibition may adversely affect host gene expression, leading to cellular toxicities. Potent inhibitors selective for HDACs that maintain LTR repression could be ideal antilatency therapeutics.
Methods: We investigated the ability of selective HDAC inhibitors to de-repress the HIV-1 LTR in both a cell line model of latency and in resting CD4 T cells isolated from patients who were aviremic on antiretroviral therapy (ART).
Results: We found that inhibition of class I HDACs increased acetylation of histones at the LTR, but that LTR chromatin was unaffected by class II HDAC inhibitors. In a latently infected cell line, inhibitors selective for class I HDACs were more efficient activators of the LTR than inhibitors that target class II HDACs. Class I HDAC inhibitors were strikingly efficient inducers of virus outgrowth from resting CD4 T cells of aviremic patients, whereas HIV was rarely recovered from patient's cells exposed to class II HDAC inhibitors.
Conclusions: Further development of selective HDAC inhibitors as part of a clinical strategy to target persistent HIV infection is warranted.
2009 Wolters Kluwer Health | Lippincott Williams & Wilkins
Figures
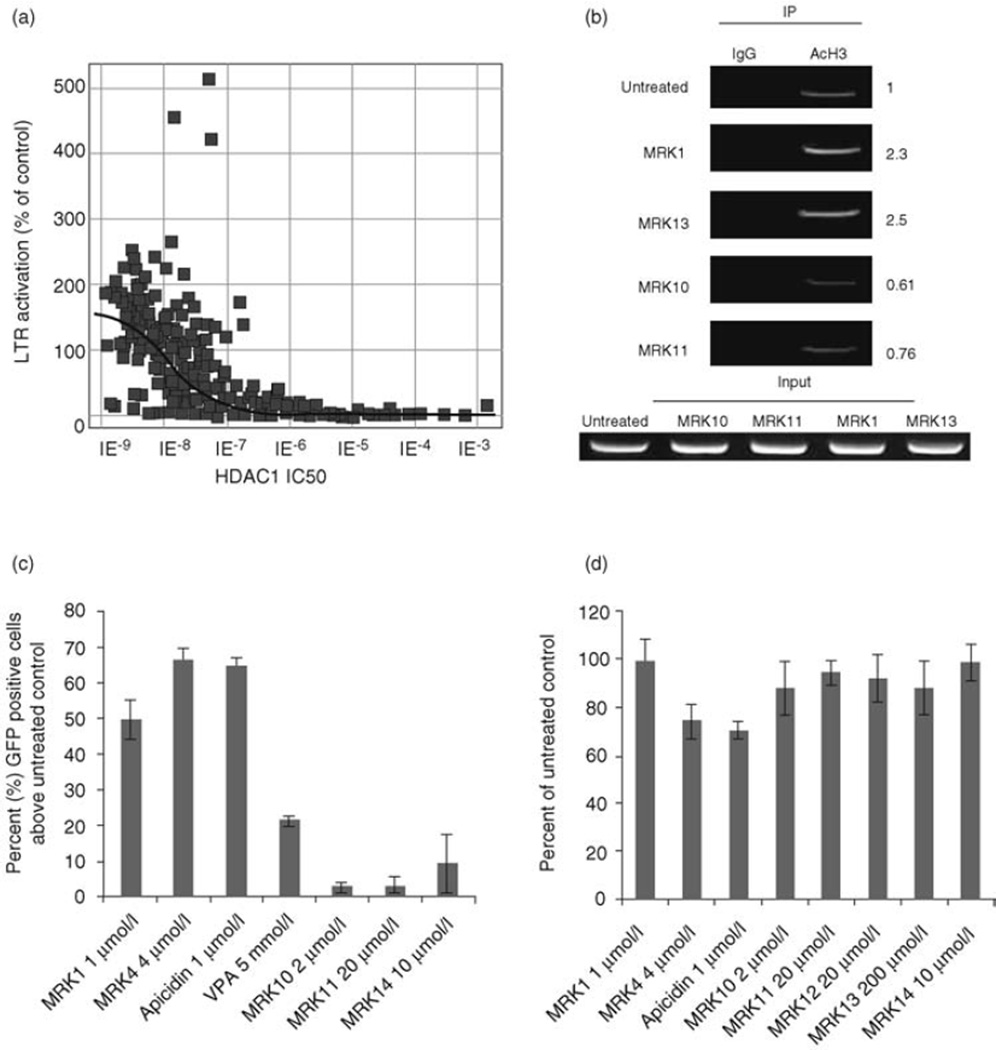


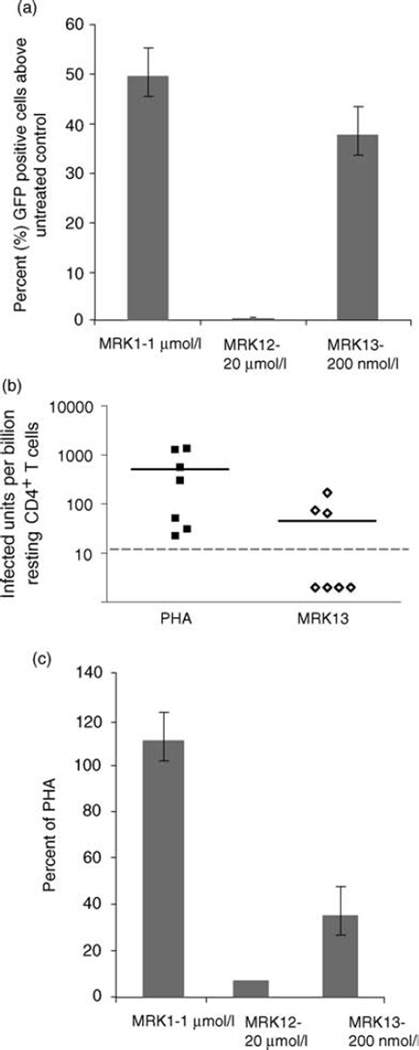
References
-
- Finzi D, Hermankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, et al. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science. 1997;278:1295–1300. - PubMed
-
- Wong JK, Hezareh M, Gunthard HF, Havlir DV, Ignacio CC, Spina CA, et al. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science. 1997;278:1291–1295. - PubMed
-
- Pazin MJ, Sheridan PL, Cannon K, Cao Z, Keck JG, Kadonaga JT, et al. NF-kappa B-mediated chromatin reconfiguration and transcriptional activation of the HIV-1 enhancer in vitro. Genes Dev. 1996;10:37–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
