

Neural stem cell transplantation benefits a monogenic neurometabolic disorder during the symptomatic phase of disease
- PMID: 19591217
- PMCID: PMC3411354
- DOI: 10.1002/stem.163
Neural stem cell transplantation benefits a monogenic neurometabolic disorder during the symptomatic phase of disease
Abstract
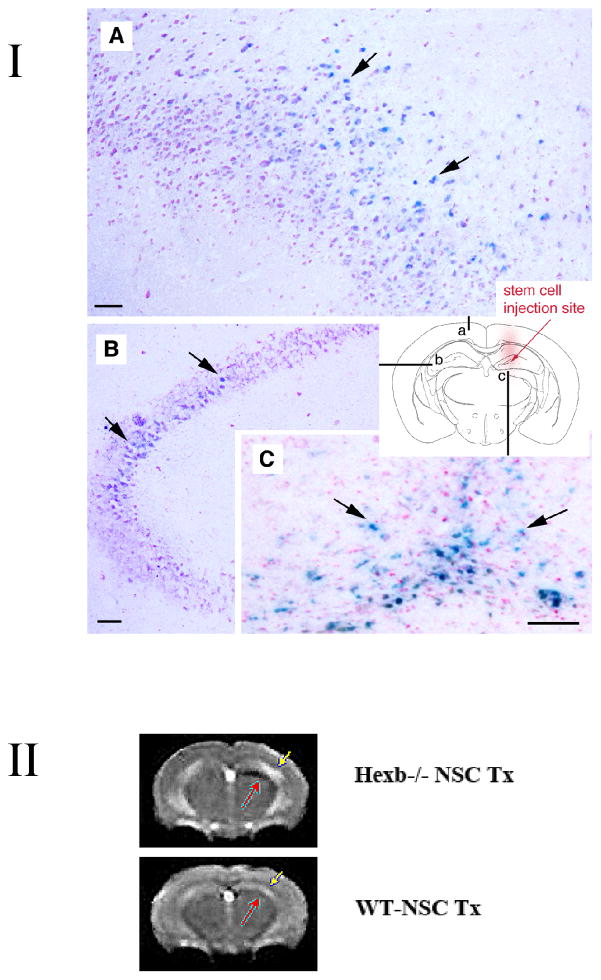
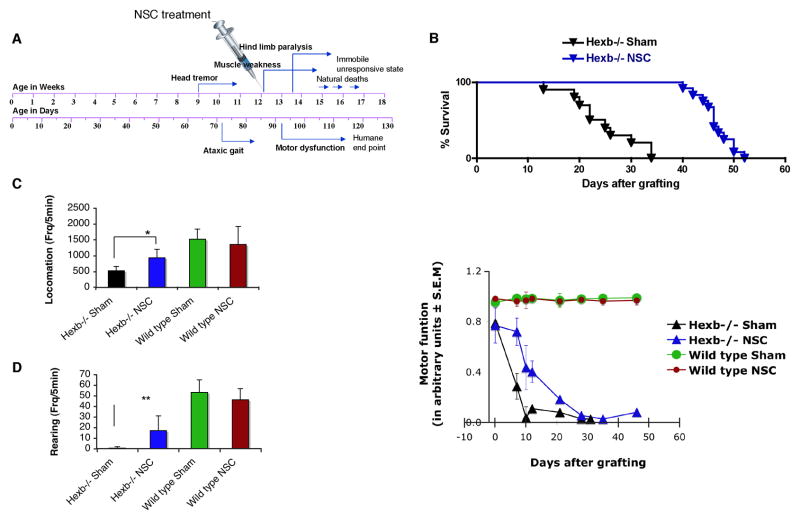
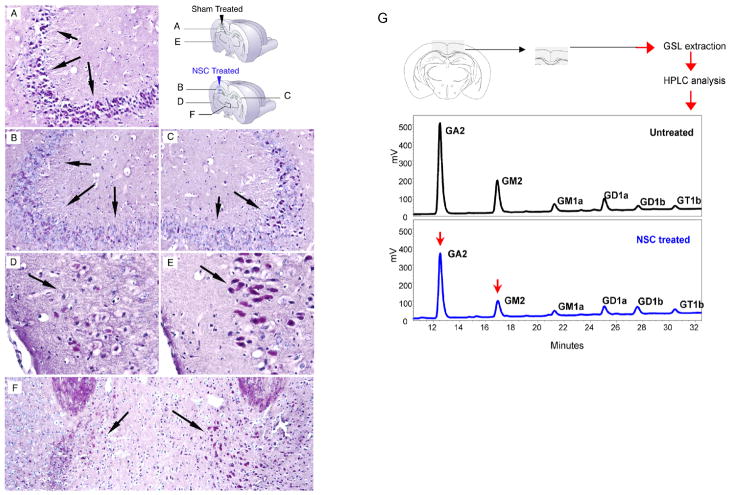
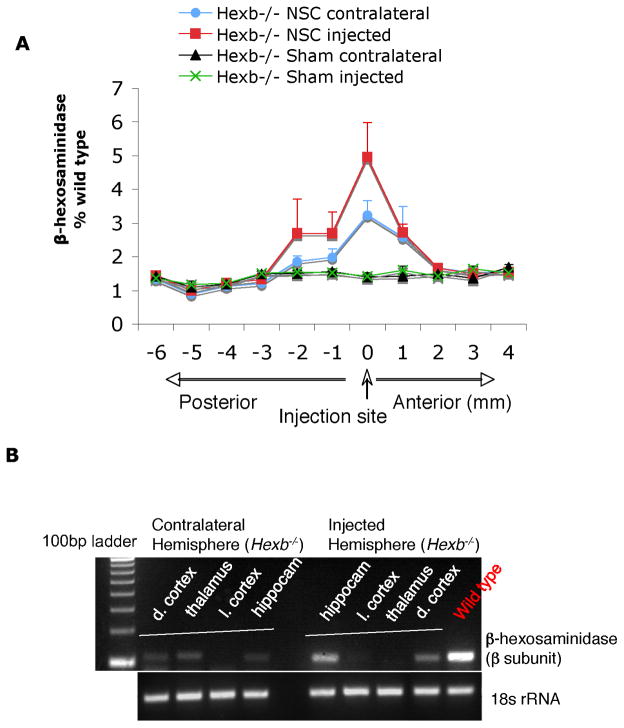
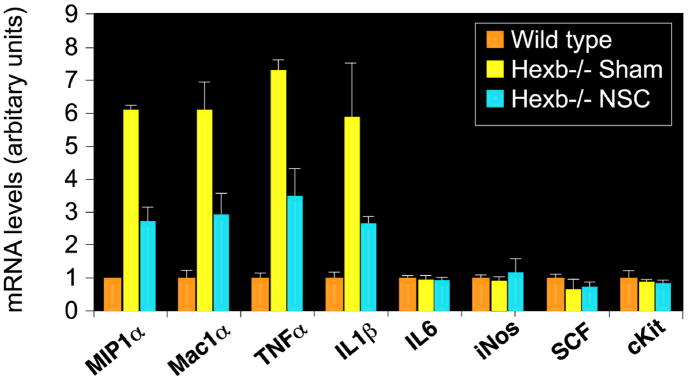
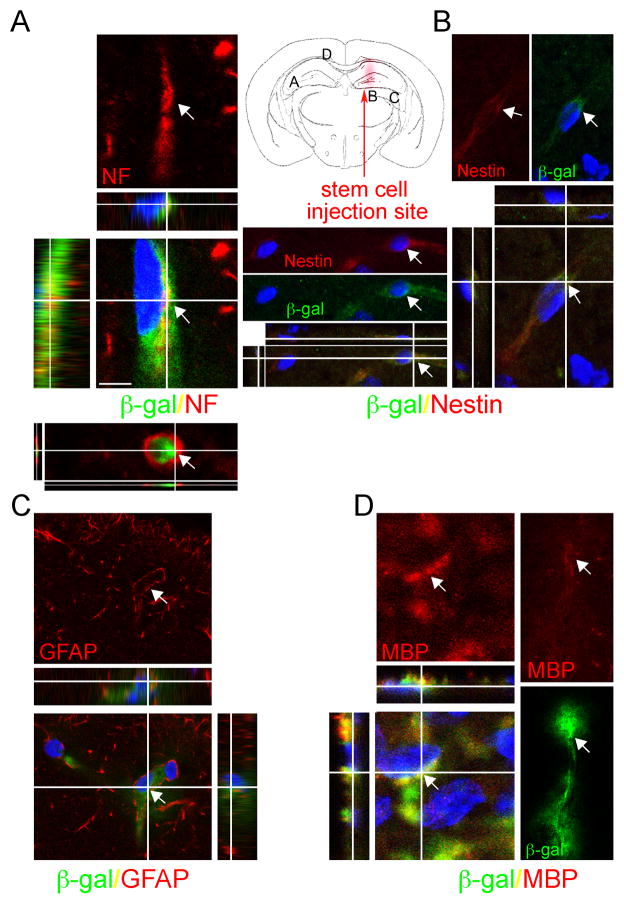
Although we and others have demonstrated that neural stem cells (NSCs) may impact such neurogenetic conditions as lysosomal storage diseases when transplanted at birth, it has remained unclear whether such interventions can impact well-established mid-stage disease, a situation often encountered clinically. Here we report that when NSCs were injected intracranially into the brain of adult symptomatic Sandhoff (Hexb(-/-)) mice, cells migrated far from the injection site and integrated into the host cytoarchitecture, restoring beta-hexosaminidase enzyme activity and promoting neuropathologic and behavioral improvement. Mouse lifespan increased, neurological function improved, and disease progression was slowed. These clinical benefits correlated with neuropathological correction at the cellular and molecular levels, reflecting the multiple potential beneficial actions of stem cells, including enzyme cross-correction, cell replacement, tropic support, and direct anti-inflammatory action. Pathotropism (i.e., migration and homing of NSCs to pathological sites) could be imaged in real time by magnetic resonance imaging. Differentially expressed chemokines might play a role in directing the migration of transplanted stem cells to sites of pathology. Significantly, the therapeutic impact of NSCs implanted in even a single location was surprisingly widespread due to both cell migration and enzyme diffusion. Because many of the beneficial actions of NSCs observed in newborn brains were recapitulated in adult brains to the benefit of Sandhoff recipients, NSC-based interventions may also be useful in symptomatic subjects with established disease.
Figures
References
-
- Wraith JE. Lysosomal disorders. Semin Neonatol. 2002;7:75–83. - PubMed
-
- Myerowitz R, Lawson D, Mizukami H, Mi Y, Tifft CJ, Proia RL. Molecular pathophysiology in Tay-Sachs and Sandhoff diseases as revealed by gene expression profiling. Hum Mol Genet. 2002;11:1343–50. - PubMed
-
- Jeyakumar M, Thomas R, Elliot-Smith E, Smith DA, van der Spoel AC, d’Azzo A, Perry VH, Butters TD, Dwek RA, Platt FM. Central nervous system inflammation is a hallmark of pathogenesis in mouse models of GM1 and GM2 gangliosidosis. Brain. 2003;126:974–87. - PubMed
-
- Sango K, Yamanaka S, Hoffmann A, Okuda Y, Grinberg A, Westphal H, McDonald MP, Crawley JN, Sandhoff K, Suzuki K, et al. Mouse models of Tay-Sachs and Sandhoff diseases differ in neurologic phenotype and ganglioside metabolism. Nat Genet. 1995;11:170–6. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
