

Srs2 disassembles Rad51 filaments by a protein-protein interaction triggering ATP turnover and dissociation of Rad51 from DNA
- PMID: 19595720
- PMCID: PMC2711036
- DOI: 10.1016/j.molcel.2009.05.026
Srs2 disassembles Rad51 filaments by a protein-protein interaction triggering ATP turnover and dissociation of Rad51 from DNA
Abstract
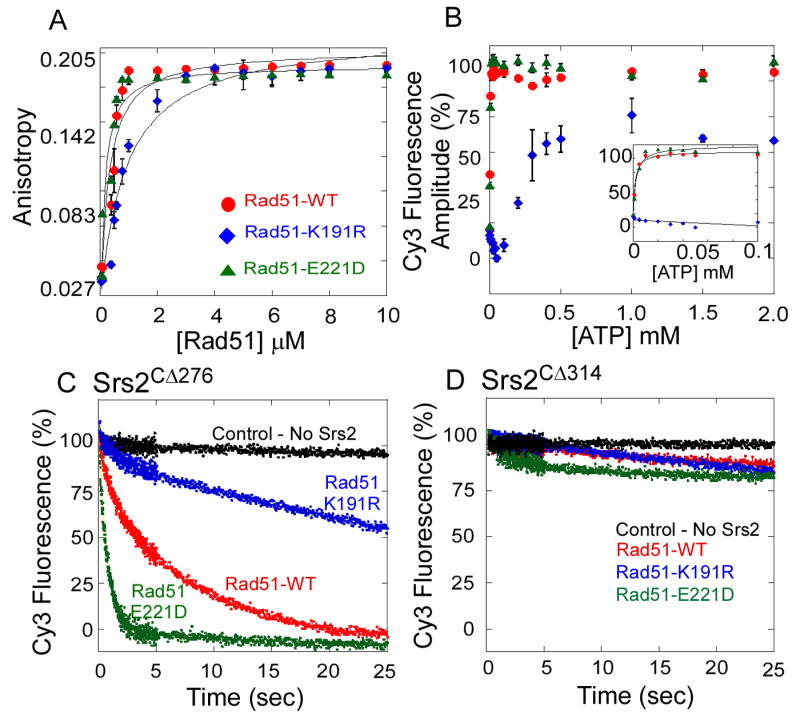
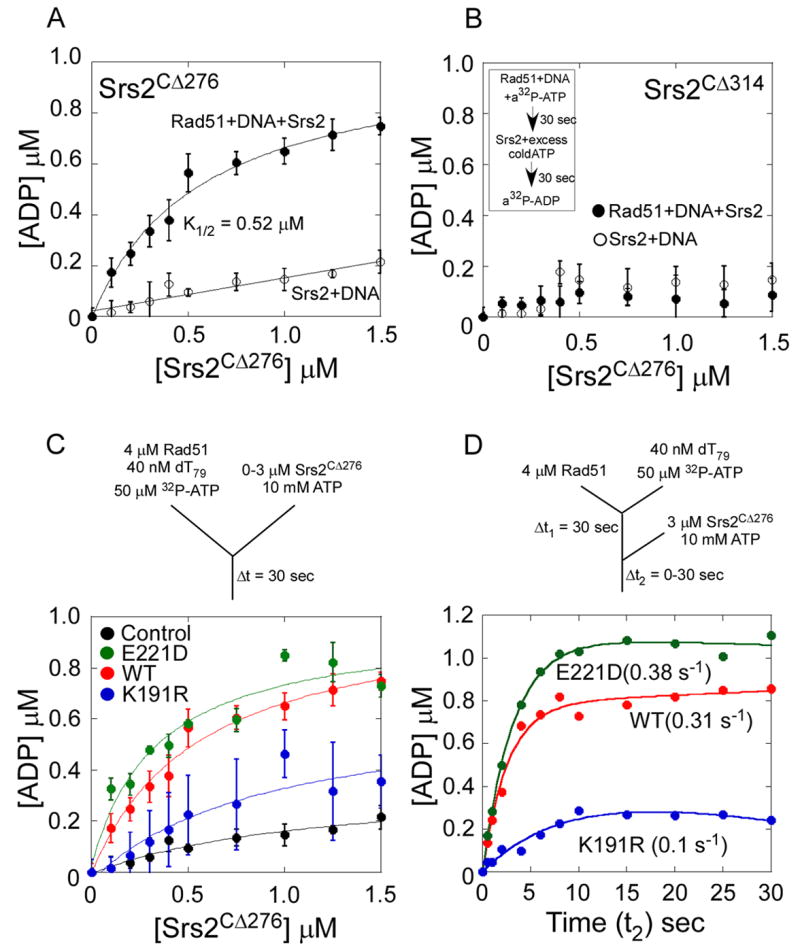
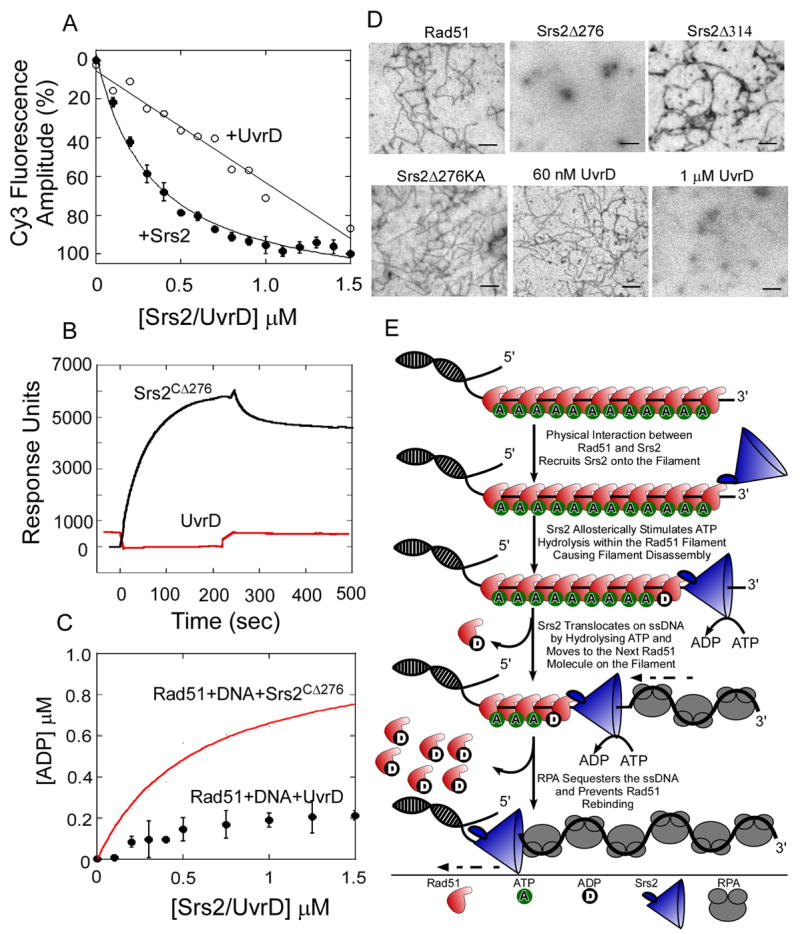
Rad51 is a DNA recombinase functioning in the repair of DNA double-strand breaks and the generation of genetic diversity by homologous recombination (HR). In the presence of ATP, Rad51 self-assembles into an extended polymer on single-stranded DNA to catalyze strand exchange. Inappropriate HR causes genomic instability, and it is normally prevented by remodeling enzymes that antagonize the activities of Rad51 nucleoprotein filaments. In yeast, the Srs2 helicase/translocase suppresses HR by clearing Rad51 polymers from single-stranded DNA. We have examined the mechanism of disassembly of Rad51 nucleoprotein filaments by Srs2 and find that a physical interaction between Rad51 and the C-terminal region of Srs2 triggers ATP hydrolysis within the Rad51 filament, causing Rad51 to dissociate from DNA. This allosteric mechanism explains the biological specialization of Srs2 as a DNA motor protein that antagonizes HR.
Figures
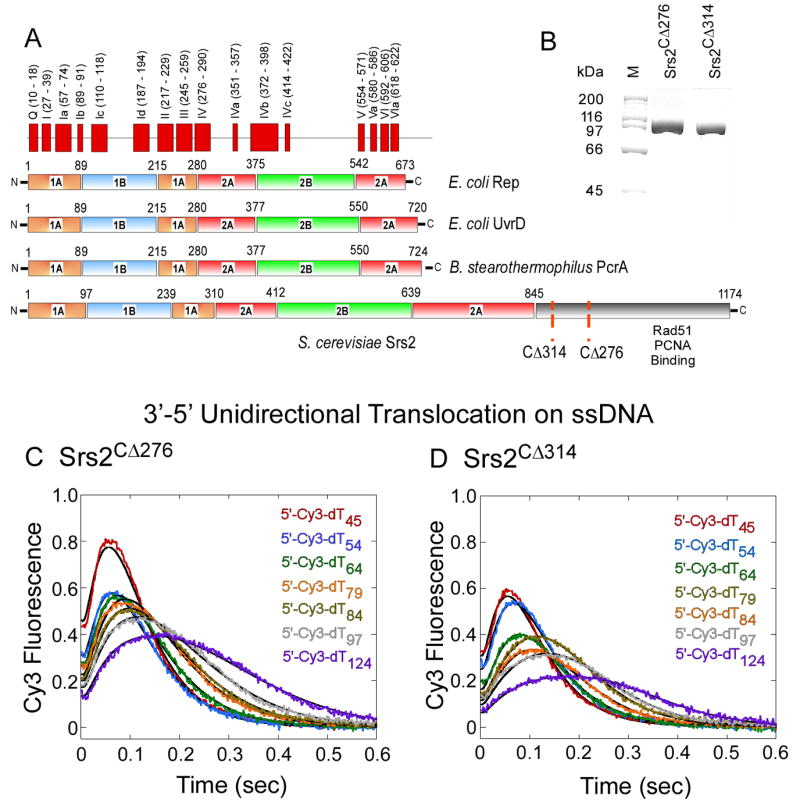
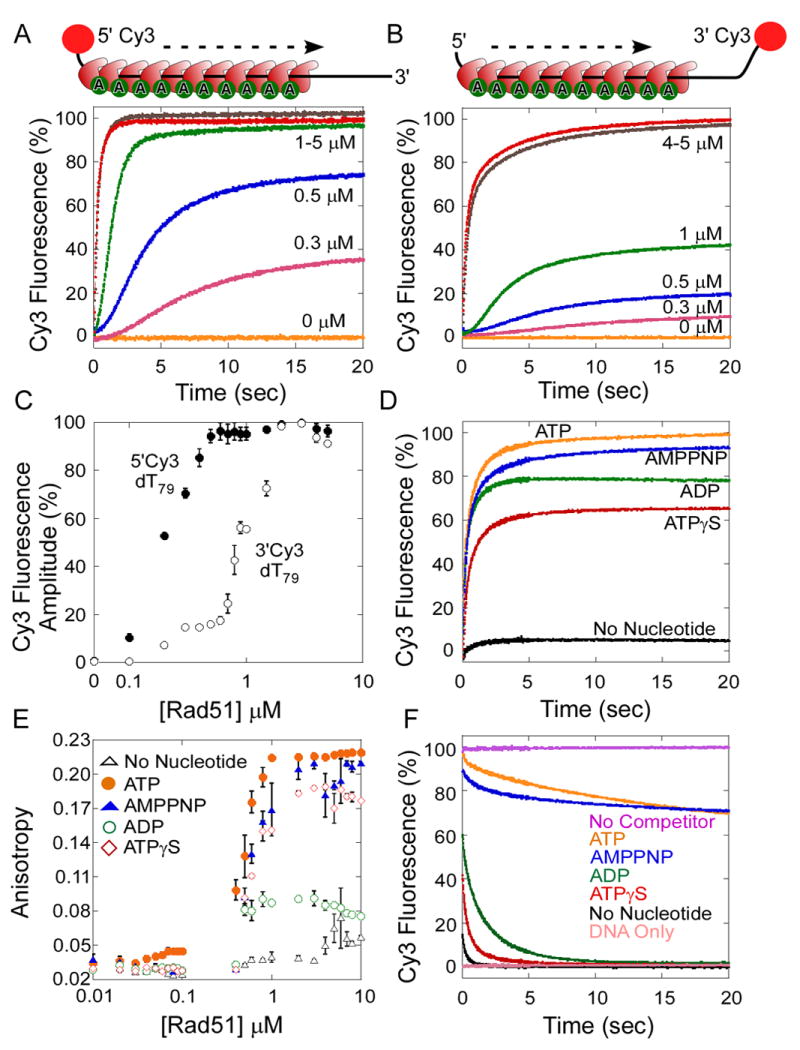
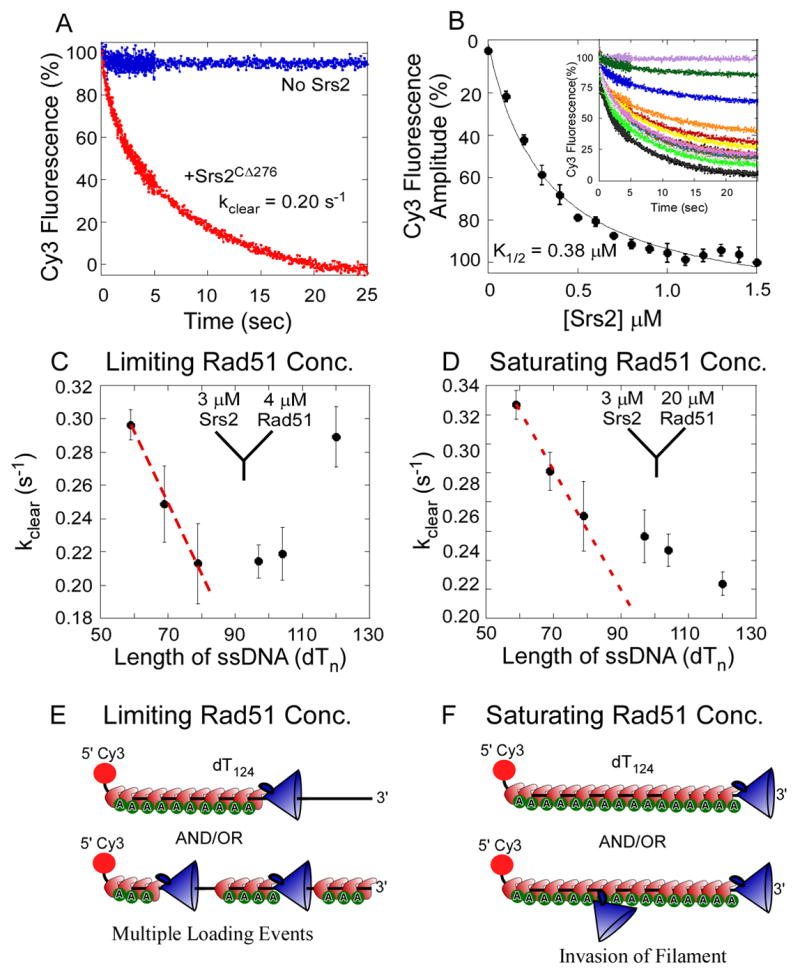
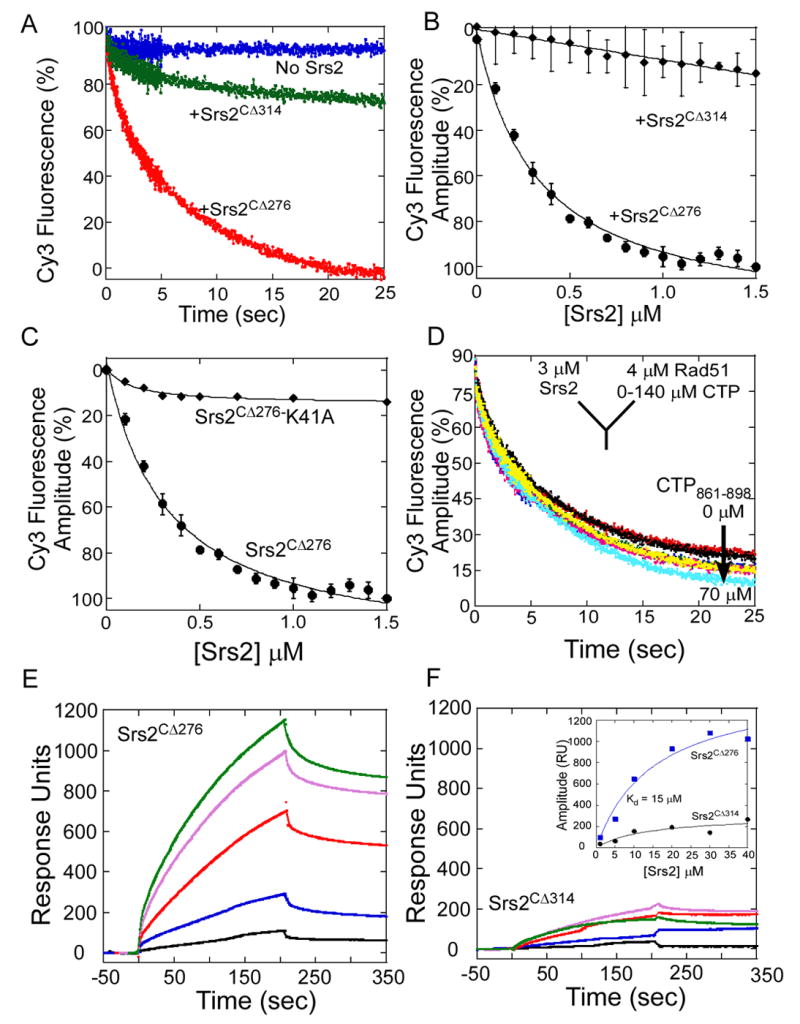
References
-
- Bianco PR, Tracy RB, Kowalczykowski SC. DNA strand exchange proteins: a biochemical and physical comparison. Front Biosci. 1998;3:D570–603. - PubMed
-
- Chi P, Van Komen S, Sehorn MG, Sigurdsson S, Sung P. Roles of ATP binding and ATP hydrolysis in human Rad51 recombinase function. DNA Repair (Amst) 2006;5:381–391. - PubMed
-
- Conway AB, Lynch TW, Zhang Y, Fortin GS, Fung CW, Symington LS, Rice PA. Crystal structure of a Rad51 filament. Nat Struct Mol Biol. 2004;11:791–796. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
