

The role of deadenylation in the degradation of unstable mRNAs in trypanosomes
- PMID: 19596809
- PMCID: PMC2760810
- DOI: 10.1093/nar/gkp571
The role of deadenylation in the degradation of unstable mRNAs in trypanosomes
Abstract
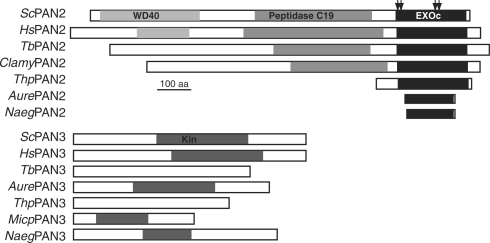
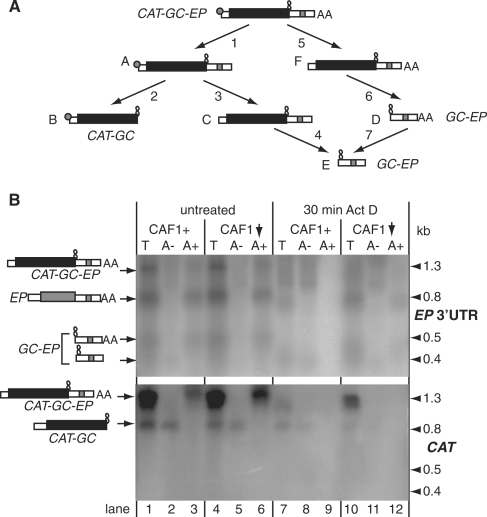
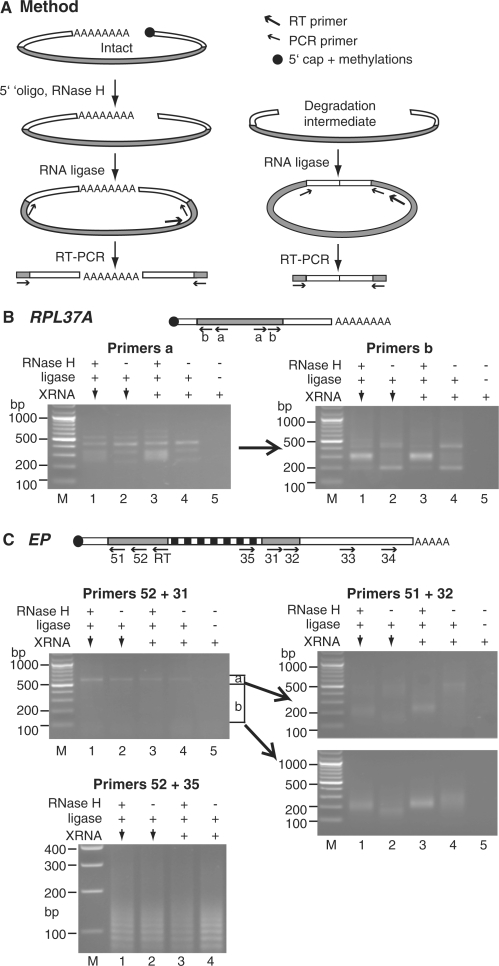
Removal of the poly(A) tail is the first step in the degradation of many eukaryotic mRNAs. In metazoans and yeast, the Ccr4/Caf1/Not complex has the predominant deadenylase activity, while the Pan2/Pan3 complex may trim poly(A) tails to the correct size, or initiate deadenylation. In trypanosomes, turnover of several constitutively-expressed or long-lived mRNAs is not affected by depletion of the 5'-3' exoribonuclease XRNA, but is almost completely inhibited by depletion of the deadenylase CAF1. In contrast, two highly unstable mRNAs, encoding EP procyclin and a phosphoglycerate kinase, PGKB, accumulate when XRNA levels are reduced. We here show that degradation of EP mRNA was partially inhibited after CAF1 depletion. RNAi-targeting trypanosome PAN2 had a mild effect on global deadenylation, and on degradation of a few mRNAs including EP. By amplifying and sequencing degradation intermediates, we demonstrated that a reduction in XRNA had no effect on degradation of a stable mRNA encoding a ribosomal protein, but caused accumulation of EP mRNA fragments that had lost substantial portions of the 5' and 3' ends. The results support a model in which trypanosome mRNAs can be degraded by at least two different, partially independent, cytoplasmic degradation pathways attacking both ends of the mRNA.
Figures





Similar articles
-
A role for Caf1 in mRNA deadenylation and decay in trypanosomes and human cells.Nucleic Acids Res. 2008 Jun;36(10):3374-88. doi: 10.1093/nar/gkn108. Epub 2008 Apr 27. Nucleic Acids Res. 2008. PMID: 18442996 Free PMC article.
-
The roles of 3'-exoribonucleases and the exosome in trypanosome mRNA degradation.RNA. 2013 Jul;19(7):937-47. doi: 10.1261/rna.038430.113. Epub 2013 May 22. RNA. 2013. PMID: 23697549 Free PMC article.
-
Trypanosome CNOT10 is essential for the integrity of the NOT deadenylase complex and for degradation of many mRNAs.Nucleic Acids Res. 2013 Jan;41(2):1211-22. doi: 10.1093/nar/gks1133. Epub 2012 Dec 4. Nucleic Acids Res. 2013. PMID: 23221646 Free PMC article.
-
mRNA deadenylation by Pan2-Pan3.Biochem Soc Trans. 2014 Feb;42(1):184-7. doi: 10.1042/BST20130211. Biochem Soc Trans. 2014. PMID: 24450649 Free PMC article. Review.
-
Roles of mRNA poly(A) tails in regulation of eukaryotic gene expression.Nat Rev Mol Cell Biol. 2022 Feb;23(2):93-106. doi: 10.1038/s41580-021-00417-y. Epub 2021 Sep 30. Nat Rev Mol Cell Biol. 2022. PMID: 34594027 Free PMC article. Review.
Cited by
-
Identification and analysis of the RNA degrading complexes and machinery of Giardia lamblia using an in silico approach.BMC Genomics. 2011 Nov 29;12:586. doi: 10.1186/1471-2164-12-586. BMC Genomics. 2011. PMID: 22126454 Free PMC article.
-
Rapid decay of unstable Leishmania mRNAs bearing a conserved retroposon signature 3'-UTR motif is initiated by a site-specific endonucleolytic cleavage without prior deadenylation.Nucleic Acids Res. 2010 Sep;38(17):5867-83. doi: 10.1093/nar/gkq349. Epub 2010 May 7. Nucleic Acids Res. 2010. PMID: 20453029 Free PMC article.
-
The Role of KH-Type Splicing Regulatory Protein (KSRP) for Immune Functions and Tumorigenesis.Cells. 2022 Apr 28;11(9):1482. doi: 10.3390/cells11091482. Cells. 2022. PMID: 35563788 Free PMC article. Review.
-
Proteomics Uncovers Novel Components of an Interactive Protein Network Supporting RNA Export in Trypanosomes.Mol Cell Proteomics. 2022 Mar;21(3):100208. doi: 10.1016/j.mcpro.2022.100208. Epub 2022 Jan 26. Mol Cell Proteomics. 2022. PMID: 35091090 Free PMC article.
-
PolyA-specific ribonuclease (PARN-1) function in stage-specific mRNA turnover in Trypanosoma brucei.Eukaryot Cell. 2011 Sep;10(9):1230-40. doi: 10.1128/EC.05097-11. Epub 2011 Jul 8. Eukaryot Cell. 2011. PMID: 21743004 Free PMC article.
References
-
- Caponigro G, Parker R. Multiple functions for the poly(A)-binding protein in mRNA decapping and deadenylation in yeast. Genes Dev. 1995;9:2421–2432. - PubMed
-
- Tharun S, Parker R. Targeting an mRNA for decapping: displacement of translation factors and association of the Lsm1p-7p complex on deadenylated yeast mRNAs. Mol. Cell. 2001;8:1075–1083. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
