

A basal ganglia-forebrain circuit in the songbird biases motor output to avoid vocal errors
- PMID: 19597157
- PMCID: PMC2709669
- DOI: 10.1073/pnas.0903214106
A basal ganglia-forebrain circuit in the songbird biases motor output to avoid vocal errors
Abstract
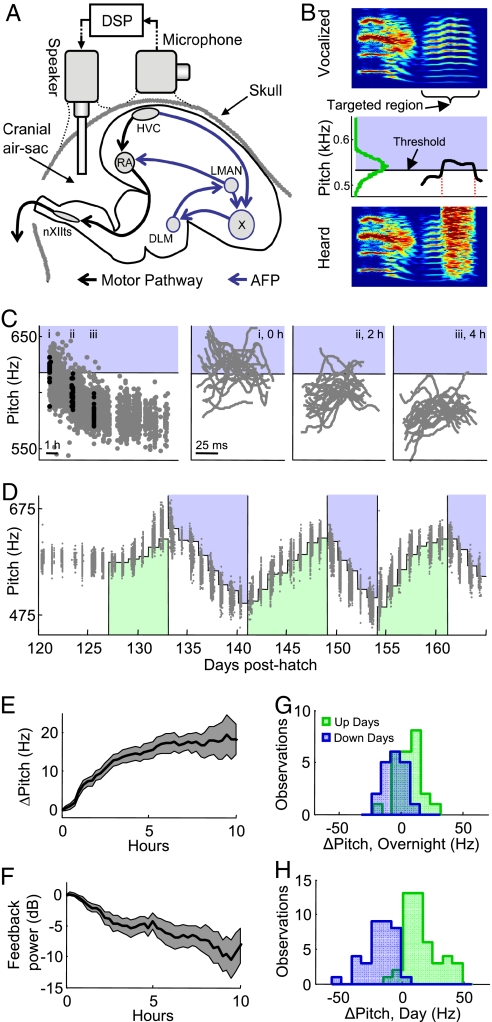
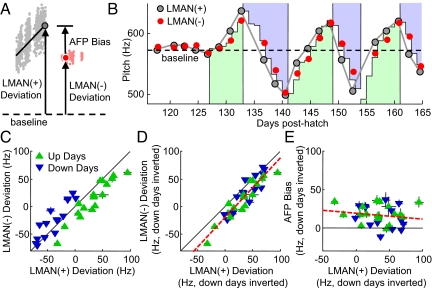
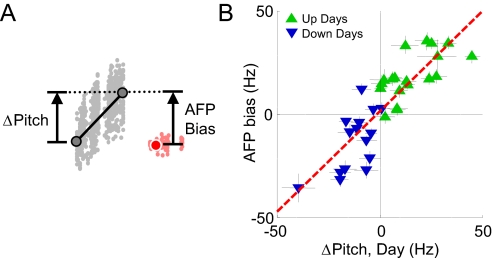
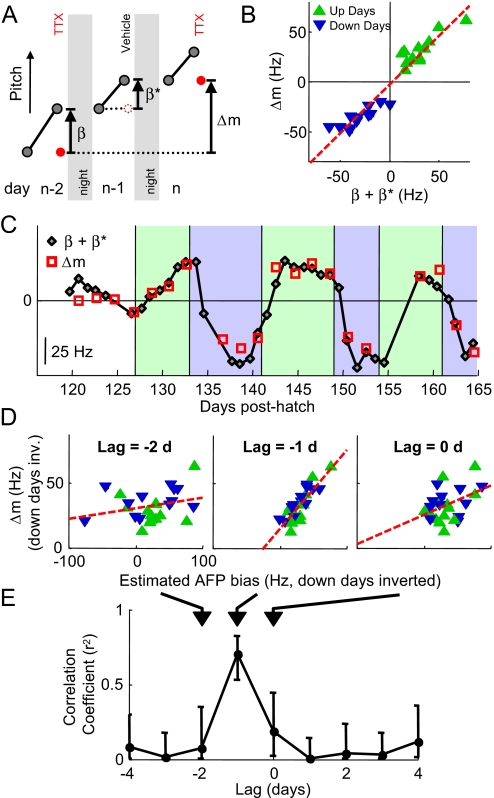
In songbirds, as in mammals, basal ganglia-forebrain circuits are necessary for the learning and production of complex motor behaviors; however, the precise role of these circuits remains unknown. It has recently been shown that a basal ganglia-forebrain circuit in the songbird, which projects directly to vocal-motor circuitry, has a premotor function driving exploration necessary for vocal learning. It has also been hypothesized that this circuit, known as the anterior forebrain pathway (AFP), may generate an instructive signal that improves performance in the motor pathway. Here, we show that the output of the AFP directly implements a motor correction that reduces vocal errors. We use disruptive auditory feedback, contingent on song pitch, to induce learned changes in song structure over the course of hours and find that reversible inactivation of the output of the AFP produces an immediate regression of these learned changes. Thus, the AFP is involved in generating an error-reducing bias, which could increase the efficiency of vocal exploration and instruct synaptic changes in the motor pathway. We also find that learned changes in the song generated by the AFP are incorporated into the motor pathway within 1 day. Our observations support a view that basal ganglia-related circuits directly implement behavioral adaptations that minimize errors and subsequently stabilize these adaptations by training premotor cortical areas.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Konishi M. The role of auditory feedback in the control of vocalization in the white-crowned sparrow. Z Tierpsychol. 1965;22:770–783. - PubMed
-
- Immelmann K. In: Bird Vocalizations. Hinde RA, editor. London: Cambridge Univ Press; 1969. pp. 61–74.
-
- Knowlton BJ, Mangels JA, Squire LR. A neostriatal habit learning system in humans. Science. 1996;273:1399–1402. - PubMed
-
- Graybiel AM. Habits, rituals, and the evaluative brain. Annu Rev Neurosci. 2008;31:359–387. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
