

CCL5-mediated endogenous antitumor immunity elicited by adoptively transferred lymphocytes and dendritic cell depletion
- PMID: 19602595
- PMCID: PMC2755640
- DOI: 10.1158/0008-5472.CAN-08-4329
CCL5-mediated endogenous antitumor immunity elicited by adoptively transferred lymphocytes and dendritic cell depletion
Abstract
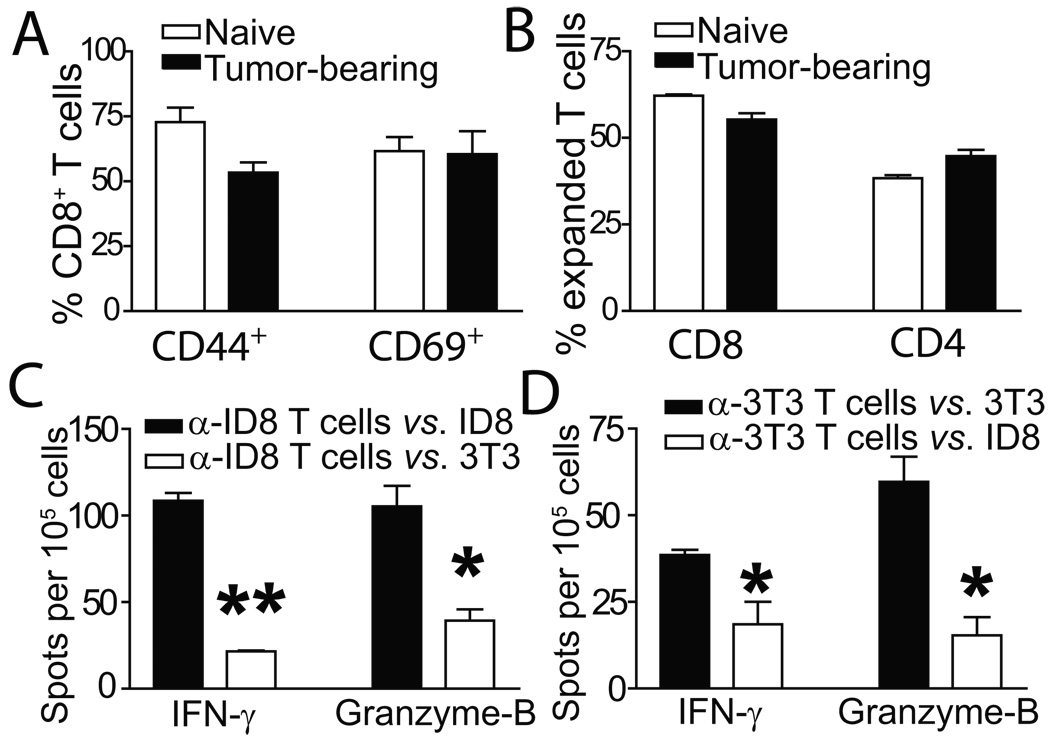
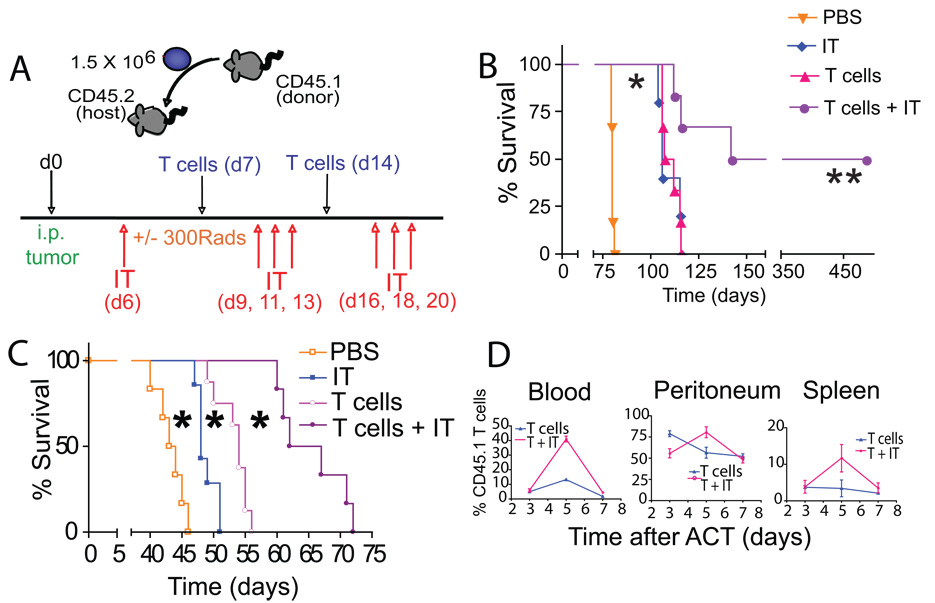
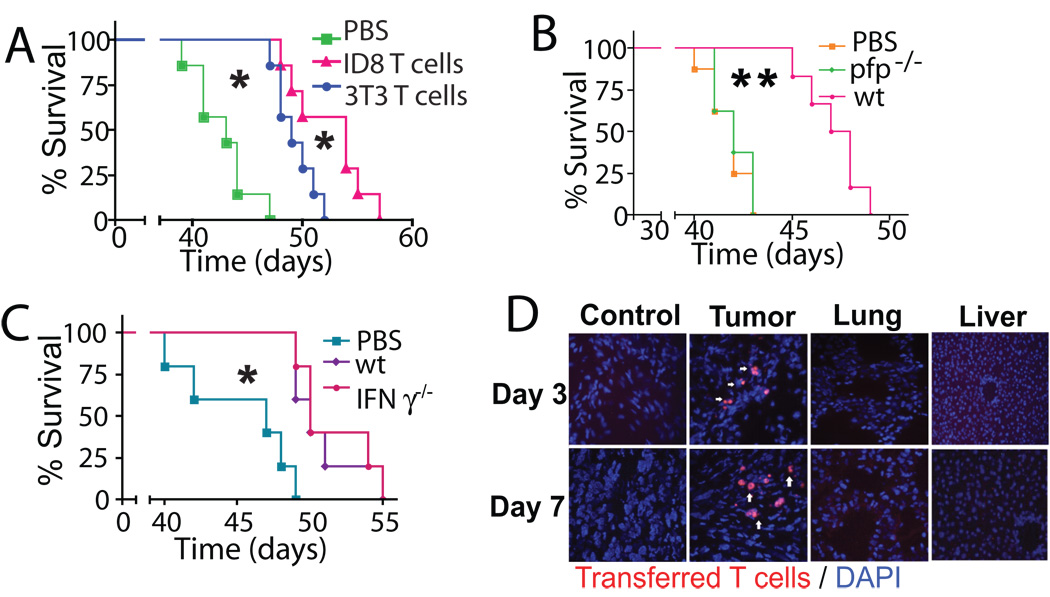
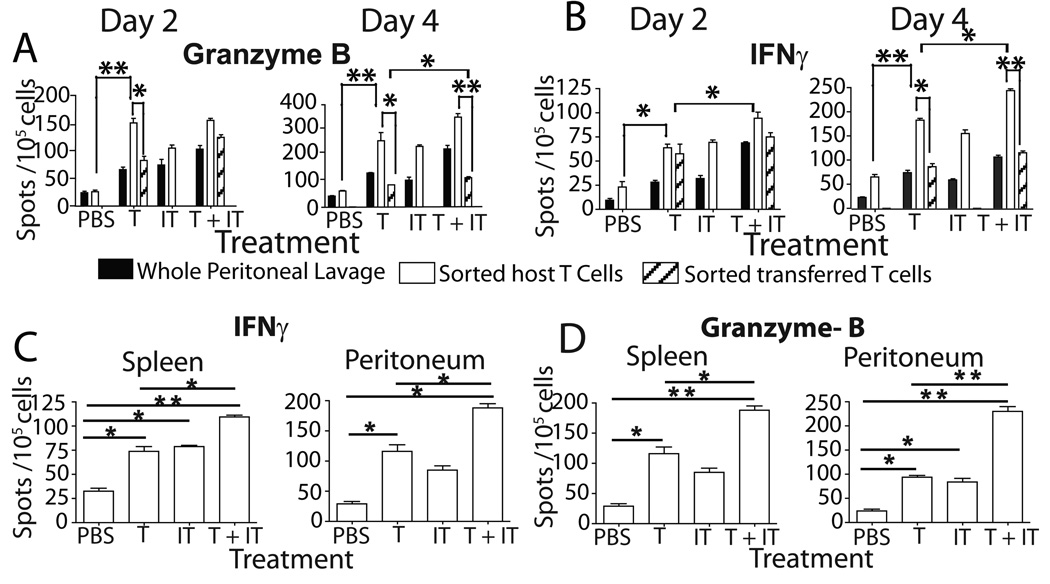
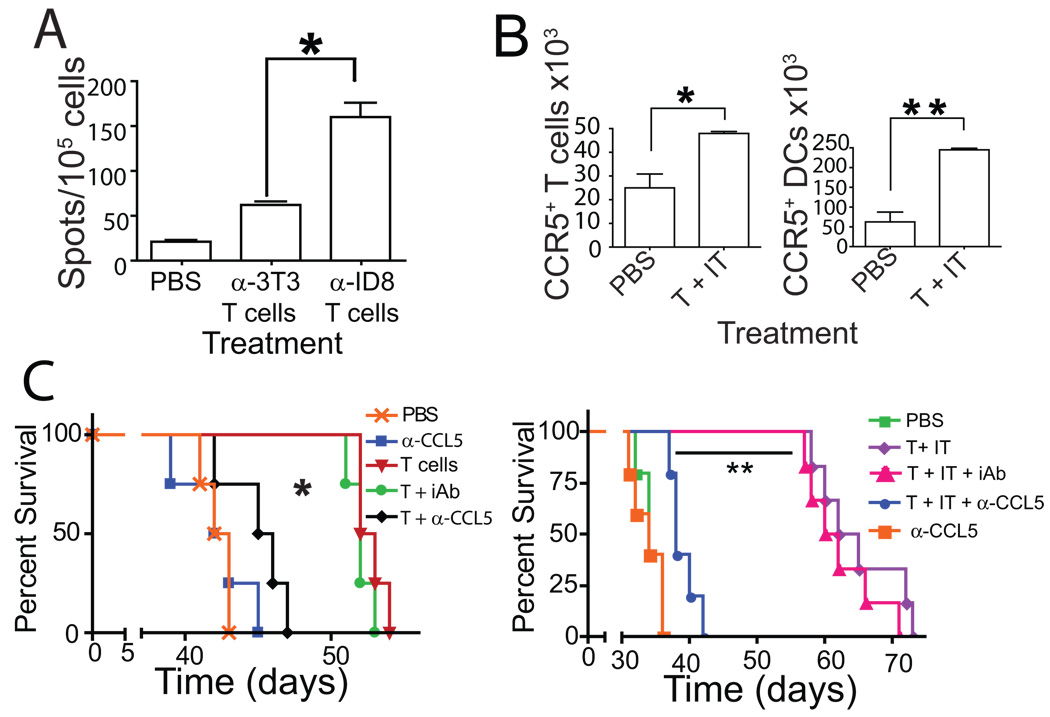
Adoptive transfer of antitumor T cells is a promisingly effective therapy for various cancers, but its effect on endogenous antitumor immune mechanisms remains largely unknown. Here, we show that the administration of naive T cells de novo primed for only 7 days against tumor antigens resulted in the durable rejection of otherwise lethal ovarian cancers when coupled with the depletion of tumor-associated immunosuppressive dendritic cells (DC). Therapeutic activity required tumor antigen specificity and perforin expression by the adoptively transferred T cells, but not IFN-gamma production. Importantly, these shortly primed T cells secreted large amounts of CCL5, which was required for their therapeutic benefit. Accordingly, transferred T cells recruited CCR5(+) DCs into the tumor, where they showed distinct immunostimulatory attributes. Activated CCR5(+) host T cells with antitumor activity also accumulated at tumor locations, and endogenous tumor-specific memory T cells remained elevated after the disappearance of transferred lymphocytes. Therefore, persistent, long-lived antitumor immunity was triggered by the administration of ex vivo activated T cells, but was directly mediated by immune cells of host origin. Our data unveil a CCL5-dependent mechanism of awakening endogenous antitumor immunity triggered by ex vivo expanded T cells, which is augmented by tumor-specific targeting of the cancer microenvironment.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
