

Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors
- PMID: 19607790
- PMCID: PMC2751885
- DOI: 10.1016/j.neuron.2009.06.014
Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors
Abstract
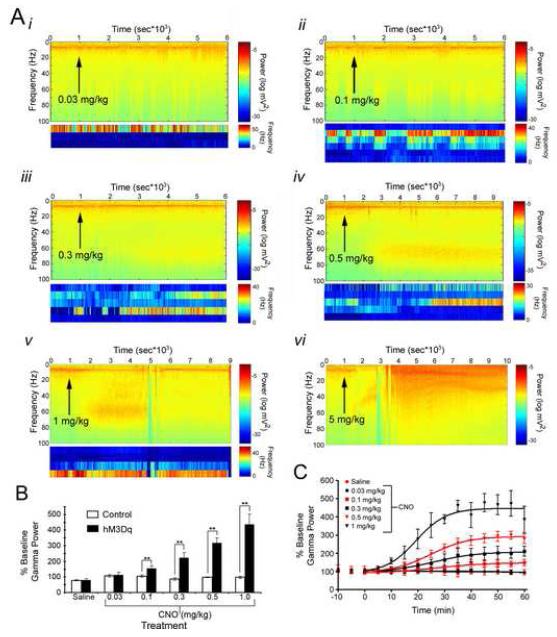
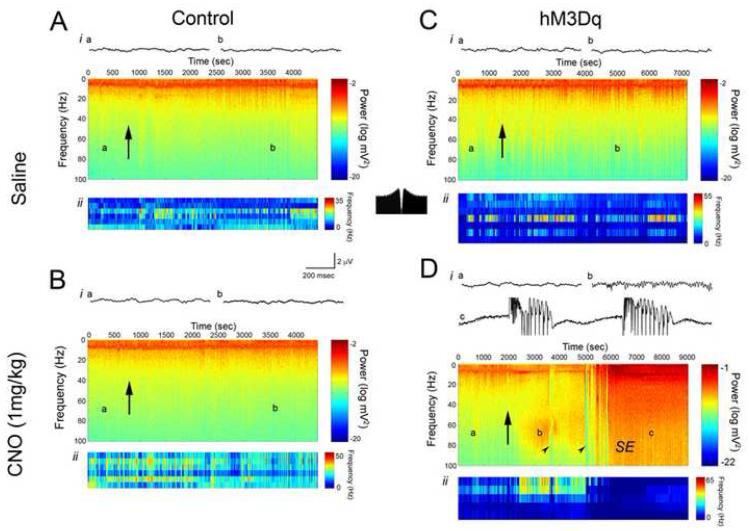
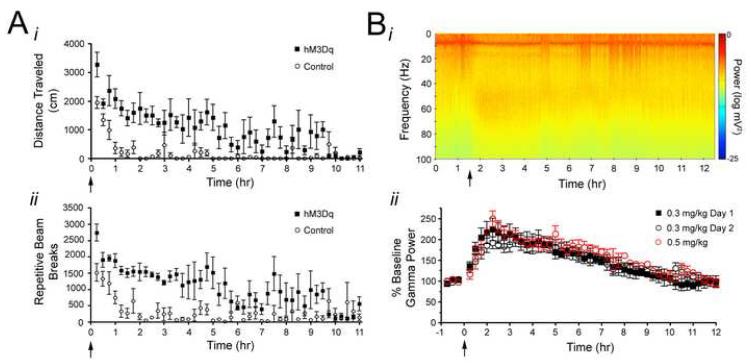
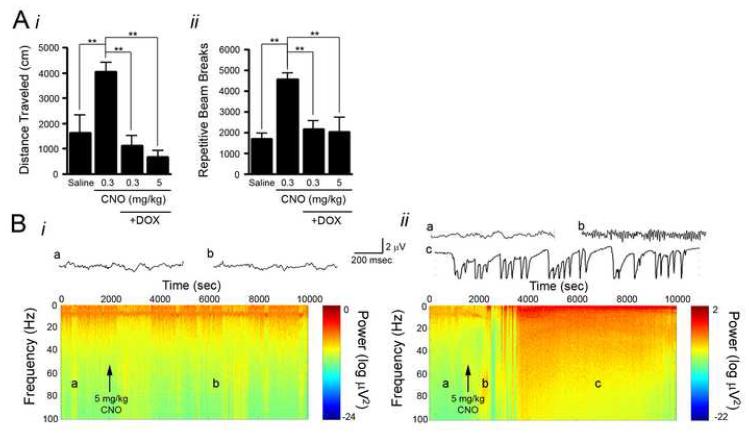
Examining the behavioral consequences of selective CNS neuronal activation is a powerful tool for elucidating mammalian brain function in health and disease. Newly developed genetic, pharmacological, and optical tools allow activation of neurons with exquisite spatiotemporal resolution; however, the inaccessibility to light of widely distributed neuronal populations and the invasiveness required for activation by light or infused ligands limit the utility of these methods. To overcome these barriers, we created transgenic mice expressing an evolved G protein-coupled receptor (hM3Dq) selectively activated by the pharmacologically inert, orally bioavailable drug clozapine-N-oxide (CNO). Here, we expressed hM3Dq in forebrain principal neurons. Local field potential and single-neuron recordings revealed that peripheral administration of CNO activated hippocampal neurons selectively in hM3Dq-expressing mice. Behavioral correlates of neuronal activation included increased locomotion, stereotypy, and limbic seizures. These results demonstrate a powerful chemical-genetic tool for remotely controlling the activity of discrete populations of neurons in vivo.
Figures
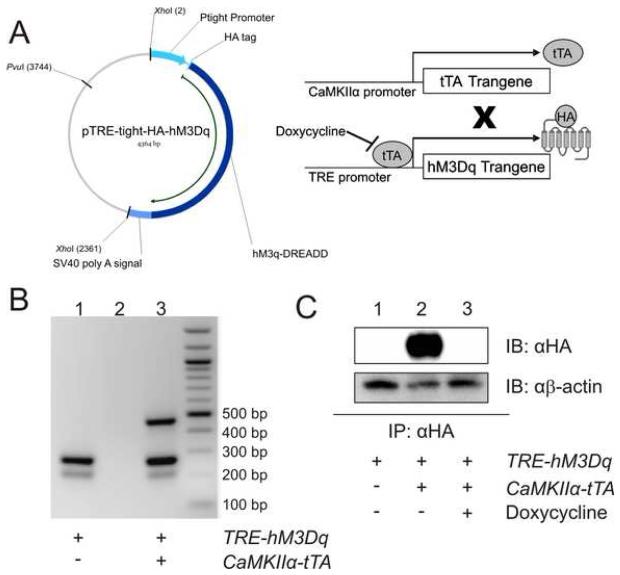
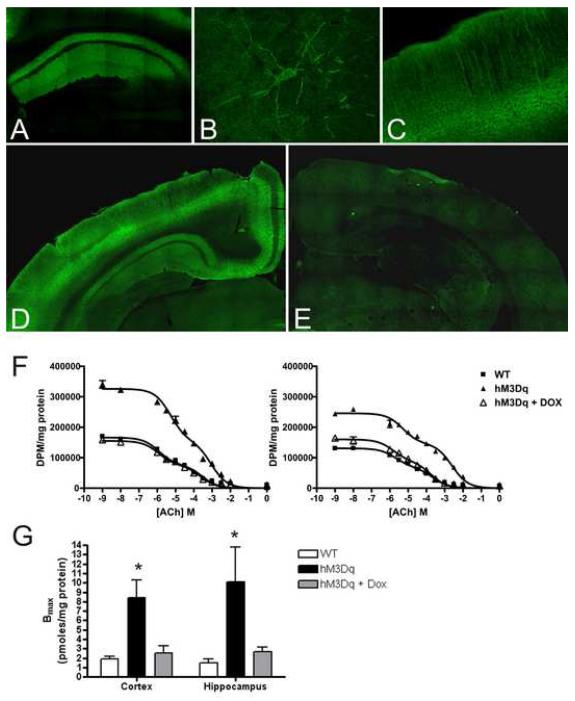
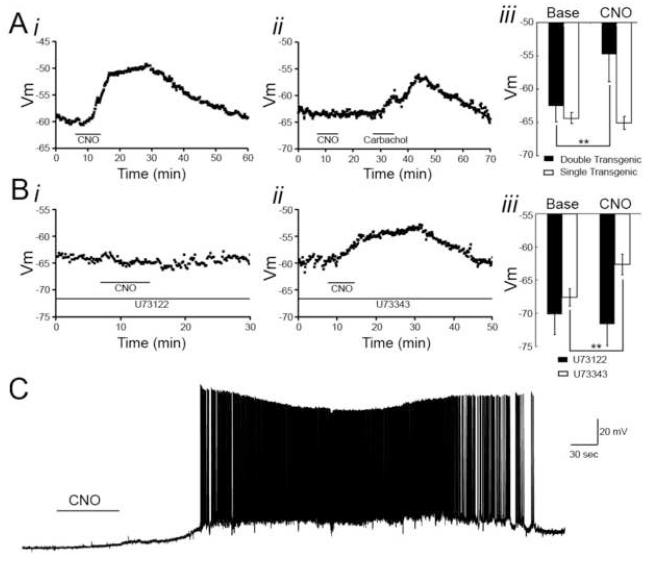
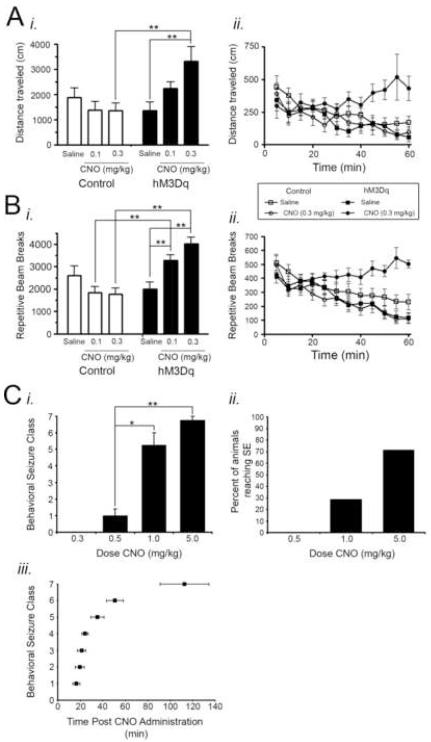
References
-
- Airan RD, Thompson KR, Fenno LE, Bernstein H, Deisseroth K. Temporally precise in vivo control of intracellular signalling. Nature. 2009;458:1025–1029. - PubMed
-
- Bender D, Holschbach M, Stocklin G. Synthesis of n.c.a. carbon-11 labelled clozapine and its major metabolite clozapine-N-oxide and comparison of their biodistribution in mice. Nucl Med Biol. 1994;21:921–925. - PubMed
-
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat Rev Neurosci. 2007;8:45–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32 NS060326/NS/NINDS NIH HHS/United States
- U19 MH082441/MH/NIMH NIH HHS/United States
- F32 GM074554/GM/NIGMS NIH HHS/United States
- T32 GM007040/GM/NIGMS NIH HHS/United States
- HD040127/HD/NICHD NIH HHS/United States
- HD03110/HD/NICHD NIH HHS/United States
- P30 HD003110/HD/NICHD NIH HHS/United States
- T32 HD040127/HD/NICHD NIH HHS/United States
- R33 NS049534/NS/NINDS NIH HHS/United States
- T32 GM007250/GM/NIGMS NIH HHS/United States
- GM007250/GM/NIGMS NIH HHS/United States
- NS049534/NS/NINDS NIH HHS/United States
- R01 NS056217/NS/NINDS NIH HHS/United States
- GM008719/GM/NIGMS NIH HHS/United States
- NS056217/NS/NINDS NIH HHS/United States
- R21 NS049534/NS/NINDS NIH HHS/United States
- MH082441-02/MH/NIMH NIH HHS/United States
- T32 GM008719/GM/NIGMS NIH HHS/United States
- GM07040-34/GM/NIGMS NIH HHS/United States
- NS060326/NS/NINDS NIH HHS/United States
- GM074554/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
