

RIG-I-dependent sensing of poly(dA:dT) through the induction of an RNA polymerase III-transcribed RNA intermediate
- PMID: 19609254
- PMCID: PMC3878616
- DOI: 10.1038/ni.1779
RIG-I-dependent sensing of poly(dA:dT) through the induction of an RNA polymerase III-transcribed RNA intermediate
Abstract
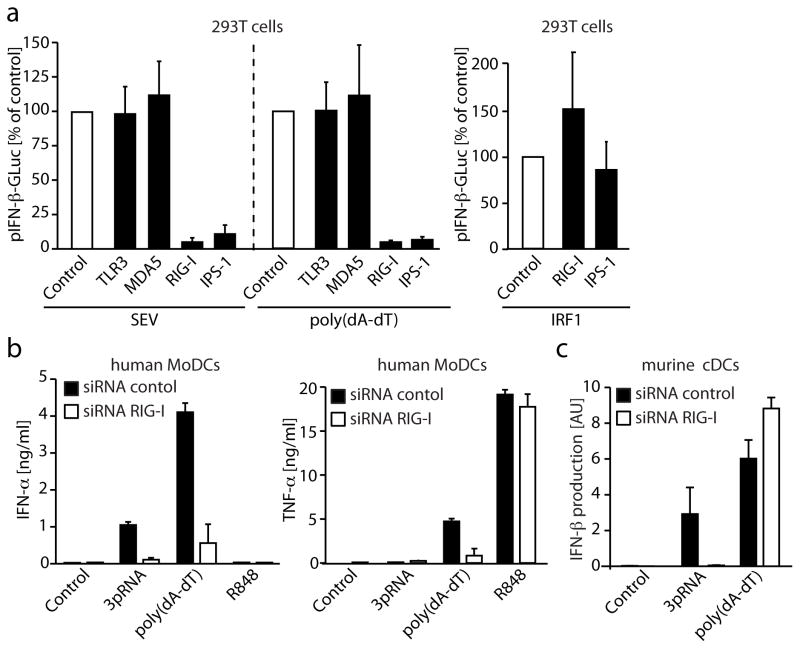
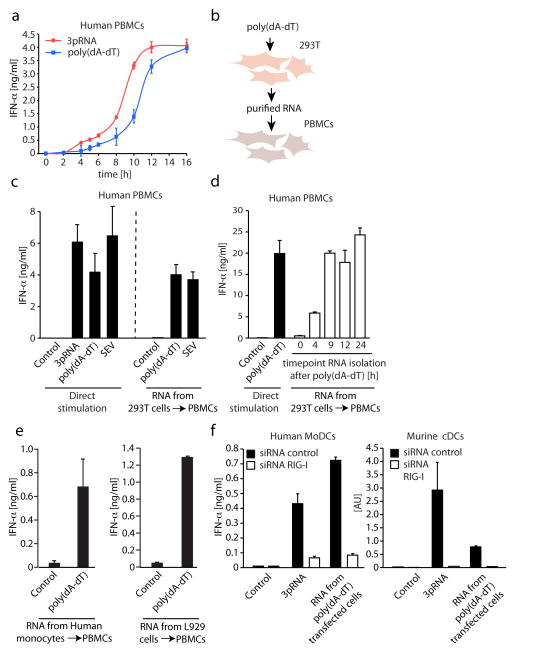
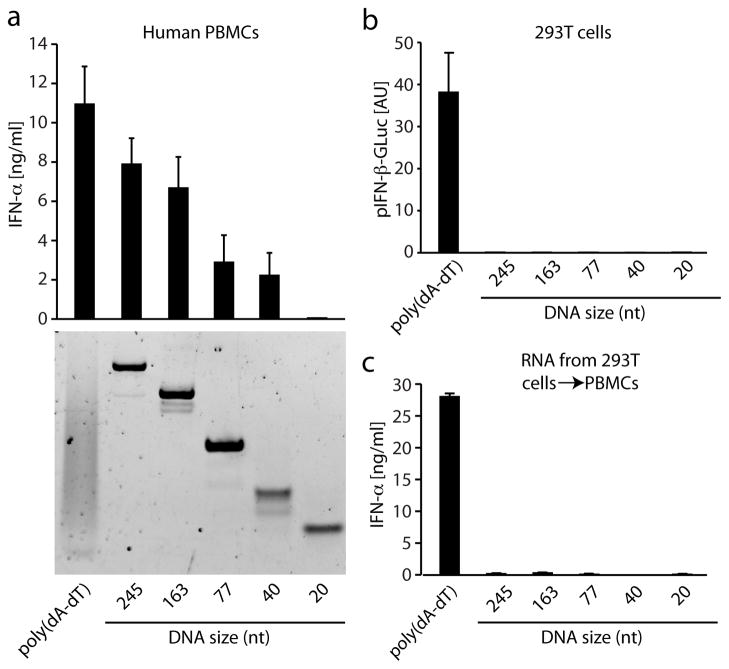
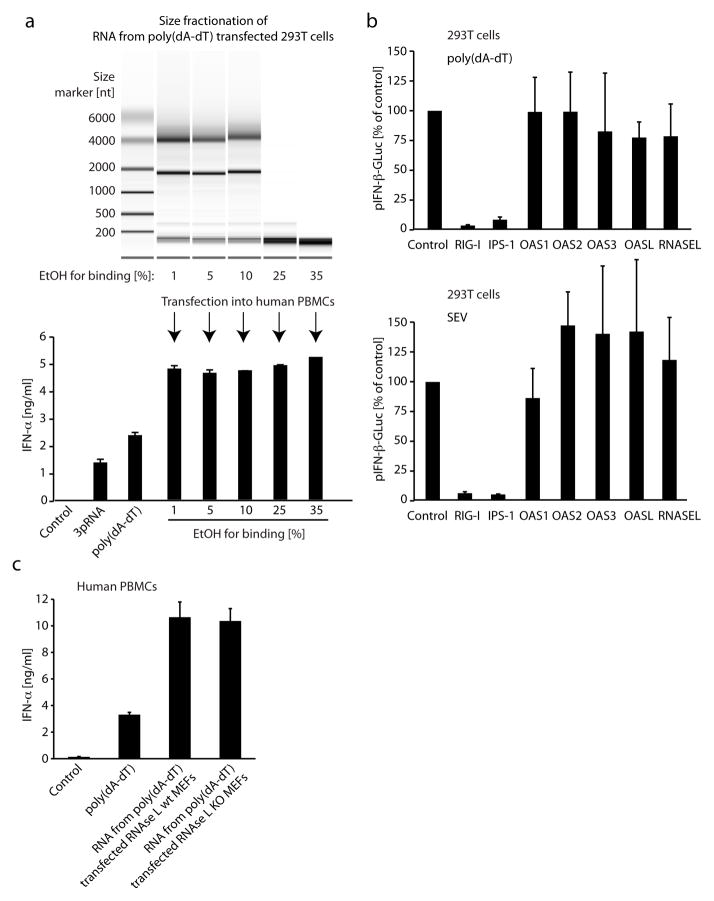
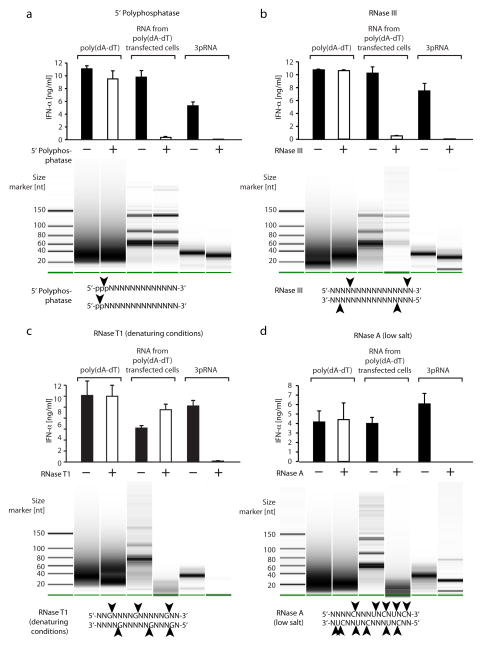
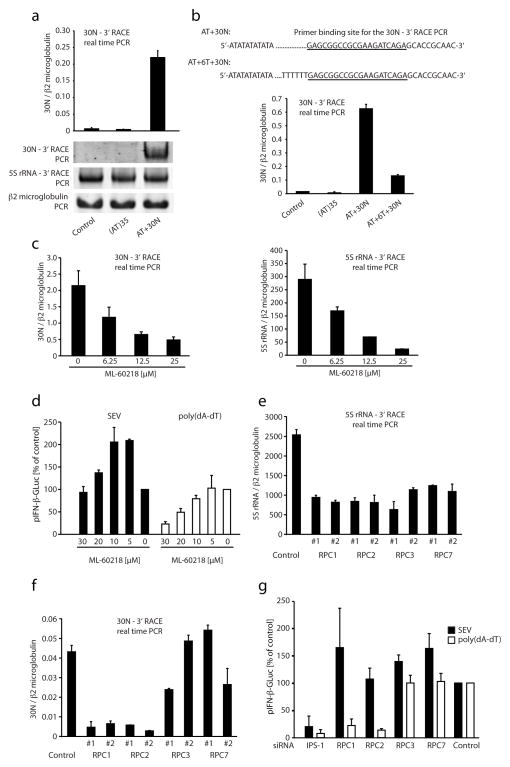
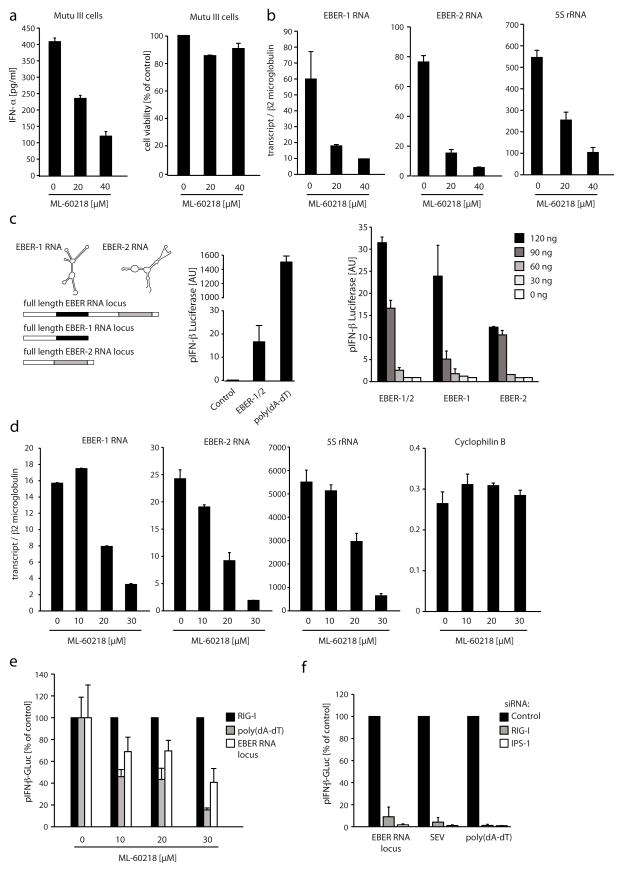
RNA is sensed by Toll-like receptor 7 (TLR7) and TLR8 or by the RNA helicases LGP2, Mda5 and RIG-I to trigger antiviral responses. Much less is known about sensors for DNA. Here we identify a novel DNA-sensing pathway involving RNA polymerase III and RIG-I. In this pathway, AT-rich double-stranded DNA (dsDNA) served as a template for RNA polymerase III and was transcribed into double-stranded RNA (dsRNA) containing a 5'-triphosphate moiety. Activation of RIG-I by this dsRNA induced production of type I interferon and activation of the transcription factor NF-kappaB. This pathway was important in the sensing of Epstein-Barr virus-encoded small RNAs, which were transcribed by RNA polymerase III and then triggered RIG-I activation. Thus, RNA polymerase III and RIG-I are pivotal in sensing viral DNA.
Figures
Comment in
-
New DNA-sensing pathway feeds RIG-I with RNA.Nat Immunol. 2009 Oct;10(10):1049-51. doi: 10.1038/ni1009-1049. Nat Immunol. 2009. PMID: 19767723 No abstract available.
References
-
- Kawai T, Akira S. TLR signaling. Semin Immunol. 2007;19:24–32. - PubMed
-
- Kawai T, Akira S. Toll-like receptor and RIG-I-like receptor signaling. Ann N Y Acad Sci. 2008;1143:1–20. - PubMed
-
- Takeda K, Akira S. Toll-like receptors in innate immunity. Int Immunol. 2005;17:1–14. - PubMed
-
- Yoneyama M, Fujita T. RIG-I family RNA helicases: cytoplasmic sensor for antiviral innate immunity. Cytokine Growth Factor Rev. 2007;18:545–551. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
