

Review
doi: 10.1038/nrmicro2182.
Staphylococcus epidermidis--the 'accidental' pathogen
Affiliations
- PMID: 19609257
- PMCID: PMC2807625
- DOI: 10.1038/nrmicro2182
Item in Clipboard
Review
Staphylococcus epidermidis--the 'accidental' pathogen
Nat Rev Microbiol.
2009 Aug.
Abstract
Although nosocomial infections by Staphylococcus epidermidis have gained much attention, this skin-colonizing bacterium has apparently evolved not to cause disease, but to maintain the commonly benign relationship with its host. Accordingly, S. epidermidis does not produce aggressive virulence determinants. Rather, factors that normally sustain the commensal lifestyle of S. epidermidis seem to give rise to additional benefits during infection. Furthermore, we are beginning to comprehend the roles of S. epidermidis in balancing the epithelial microflora and serving as a reservoir of resistance genes. In this Review, I discuss the molecular basis of the commensal and infectious lifestyles of S. epidermidis.
Figures
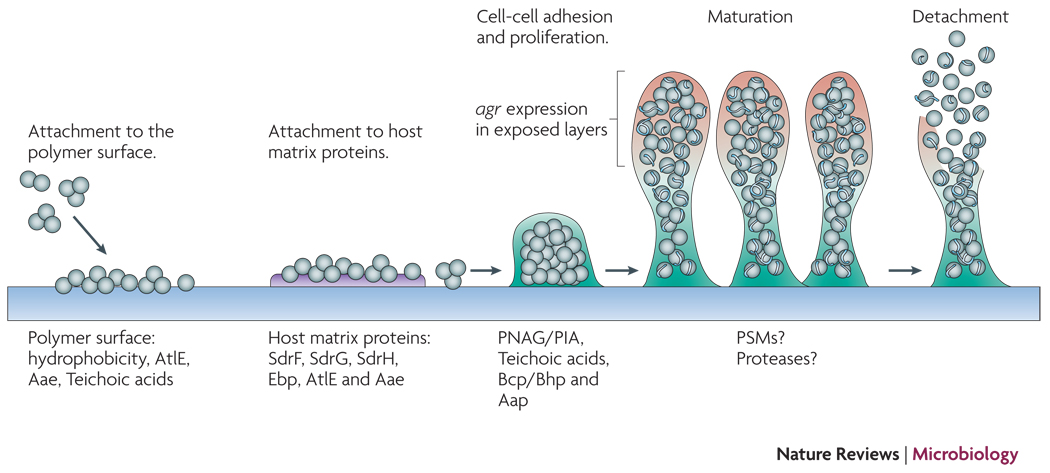
Attachment to uncoated material is mainly dependent on cell surface hydrophobicity, while dedicated surface proteins mediate adhesion to host matrix-covered devices. Afterwards, exopolysaccharide, specific proteins, and accessory macromolecules provide intercellular aggregation. Mechanisms of biofilm maturation, structuring, and detachment are poorly understood, but possibly involve quorum-sensing controlled expression of detergent-like peptides and proteolytic activity in exposed layers of the biofilm. Genome-wide gene expression is significantly different in the biofilm compared to the planktonic mode of growth and includes down-regulation of basic cell processes.
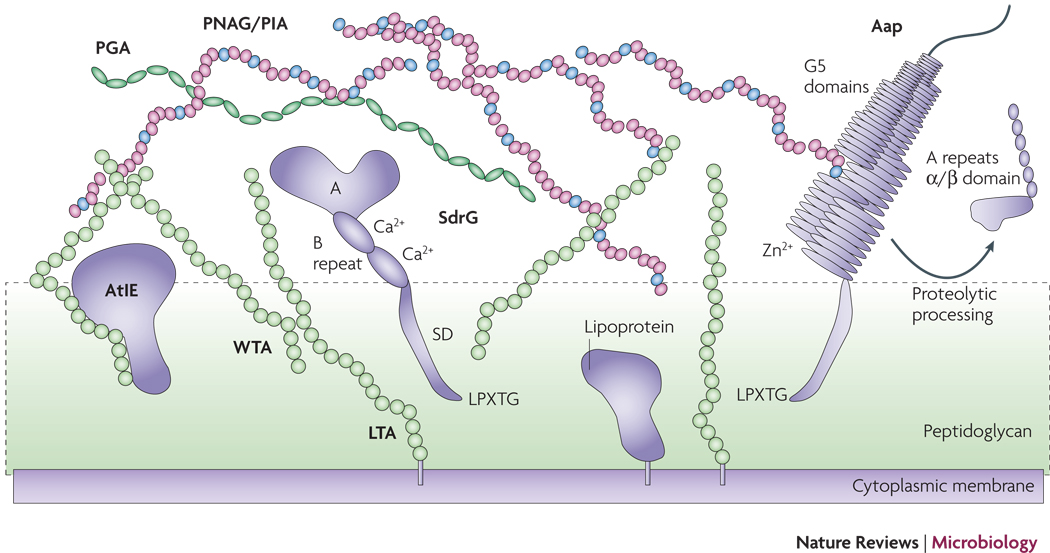
Proteins such as SdrG and Aap may be attached to the cell surface via sortase-catalyzed covalent anchoring. These proteins harbor a characteristic LPXTG motif at the C-terminus, of which the threonine residue is linked to peptidoglycan. Many autolysins such as AtlE are anchored non-covalently, likely via interaction with teichoic acids. Furthermore, lipoproteins are surface-attached via their fatty acid anchor that penetrates the cytoplasmic membrane. AtlE is a bifunctional adhesin/autolysin that contributres to biofilm formation by its surface hydrophobicity and to host matrix protein binding. SdrG is an example of the Sdr protein family of MSCRAMMs. It stretches the peptidoglycan layer by its SD repeat region and binds fibrinogen via its A region. The B repeats harbor a Ca2+ binding EF-hand domain. Aap proteins aggregate via Zn2+-dependent G5 domains and form fibrils that likely connect cells in the biofilm matrix. G5 domains also bind N-acetylglucosamine and may thus interact with the N-acetylglucosamine exopolysaccharide PNAG/PIA. PNAG/PIA is cationic and likely interacts with negatively charged surface polymers such as teichoic acids (lipoteichoic acids, LTA and wall teichoic acids, WTA) and poly-γ-glutamic acid (PGA).
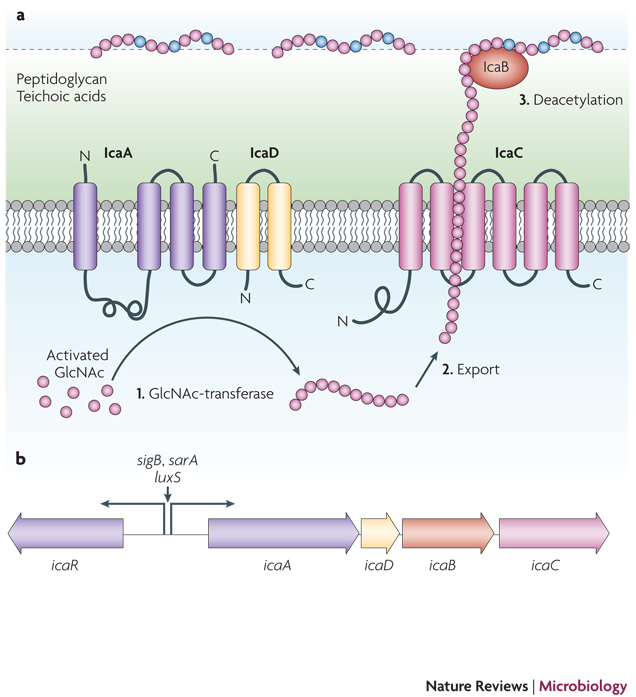
The immune evasion and biofilm aggregation exopolysaccharide PNAG/PIA, a partially de-acetylated β 1–6 linked N-acetylglucosamine homopolymer, is synthesized by the membrane-located N-acetylglucosamine transferase IcaA that needs the accessory IcaD membrane protein for activity. The growing PNAG/PIA chain is likely exported by the IcaC membrane protein. After export, the surface-located IcaB de-acetylase removes some of the N-acetyl groups, giving the polymer a cationic character that is essential for surface attachment. The Ica proteins are encoded in the ica gene locus, which contains the icaADBC operon and the icaR gene encoding a regulatory protein. Expression of the icaADBC operon is regulated either directly at the icaA promoter or via expression of IcaR by a series of global regulatory proteins. Furthermore, insertion and excision of the IS256 element may turn PNAG/PIA expression off and on.
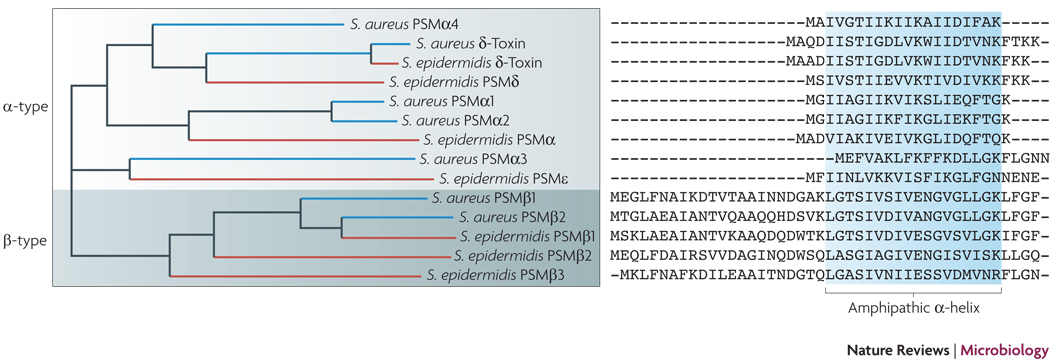
Sequence alignment of S. epidermidis and S. aureus PSMs. PSMs serve as immune evasion molecules to their bacterial producer and, on the other hand, as PAMPs for pathogen recognition to the host. All PSMs contain an amphipathic α-helix and N-terminal N-formyl methionine, as they are secreted as the direct translational product without processing in an unknown manner. PSMs of the α-type are relatively short, ~ 20–25 amino acids. Particularly the S. aureus PSMα peptides 1 through 4 are strongly cytolytic. PSMs of the β-type are longer, ~ 45 amino acids, and do not have considerable cytolytic activity. Only the δ-toxin, an α-type PSM with moderate cytolytic activity, and the β-type PSMs are secreted by S. epidermidis in large amounts. Although part of the psmβ operon, the PSMβ3 peptide is not found in S. epidermidis culture filtrates for unknown reasons. The psmβ1 gene is duplicated in some strains of S. epidermidis.
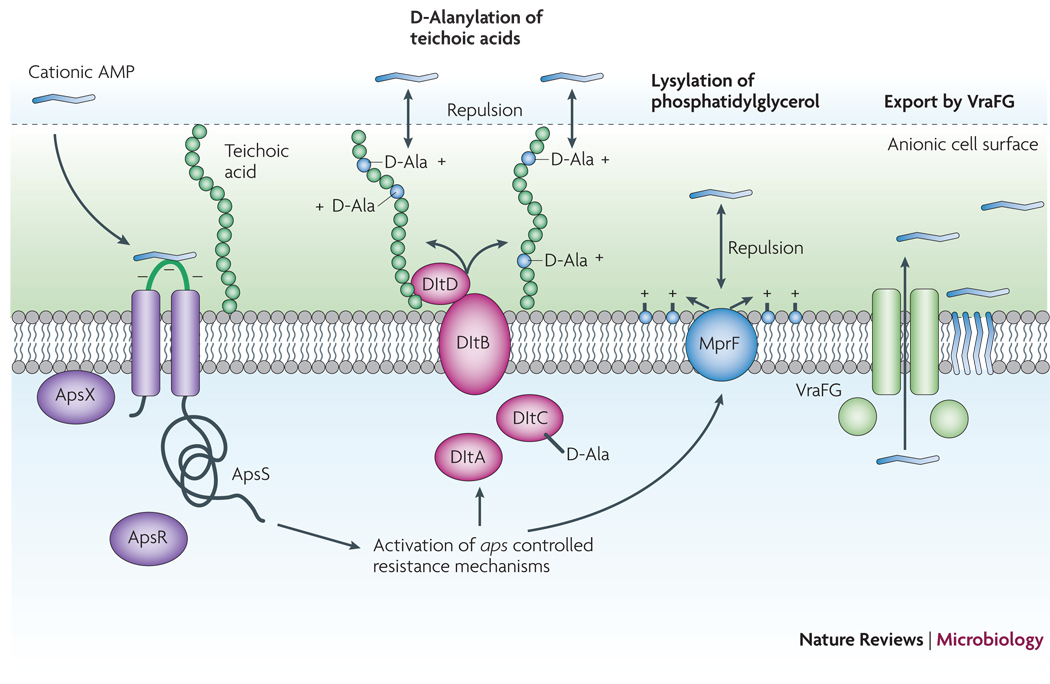
Cationic AMPs attach to the negatively charged bacterial surface and membrane by electrostatic interaction, a prerequisite for AMP antimicrobial activity, which is often based on pore formation in the bacterial cytoplasmic membrane. The S. epidermidis ApsS AMP sensor has one short extracellular loop with a high density of negatively charged amino acid residues that interacts with cationic AMPs. Transduction of this signal via ApsS and the accessory, essential ApsX, which has a yet unknown function, triggers expression of key AMP resistance mechanisms. The D-alanylation of teichoic acids, encoded by the products of the dlt operon, and lysylation of phosphatidylglycerol, catalyzed by the MprF enzyme, result in a decreased negative charge of the cell surface and membrane, respectively, leading to decreased attraction, or repulsion, of cationic AMPs. The VraFG ABC transporter also promotes resistance to AMPs and likely functions as an AMP exporter.
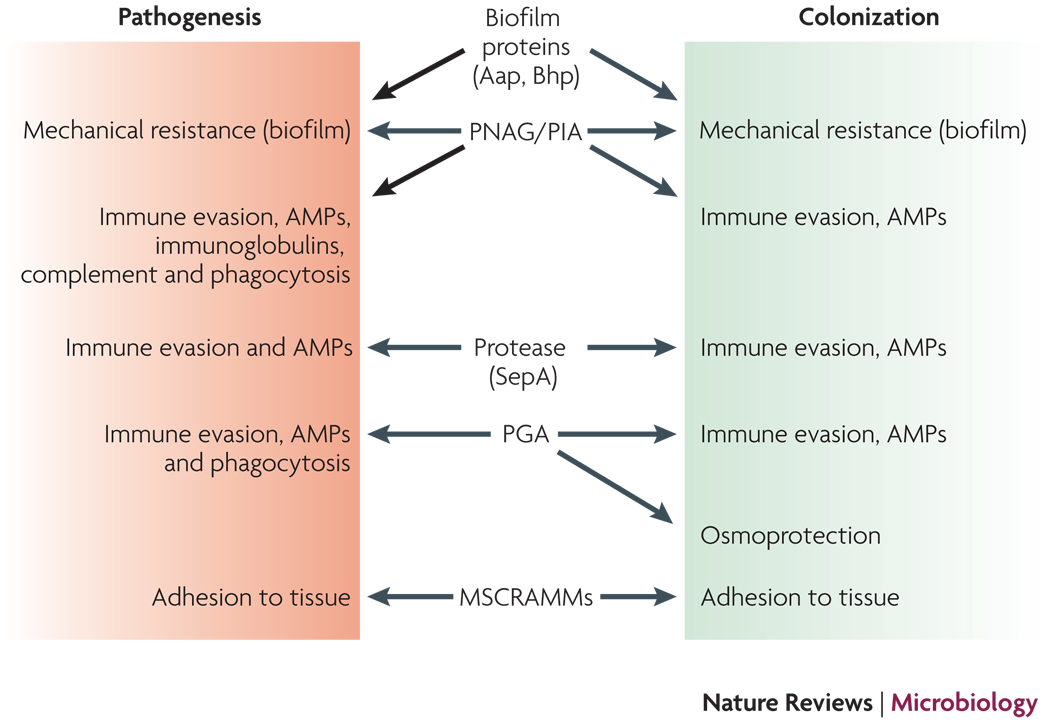
Determinants that are believed to contribute to both S. epidermidis colonization and pathogenesis are shown. In animal models, only the roles of PNAG/PIA, PGA, and the MSCRAMM SdrG in infection have been demonstrated. Other roles are based on in vitro experiments and environmental challenges during colonization and infection. Not shown are regulators such as agr or sigB that control many of the depicted determinants and may thus also have important functions during both S. epidermidis lifestyles.
References
-
- CDC. National Nosocomial Infections Surveillance (NNIS) System Report, data summary from January 1992 through June 2004, issued October 2004. Am J Infect Control. 2004;32:470–485. - PubMed
-
- Uckay I, et al. Foreign body infections due to Staphylococcus epidermidis. Ann Med. 2009;41:109–119. - PubMed
-
- Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science. 1999;284:1318–1322. - PubMed
-
- Kloos W, Schleifer KH. In: Bergey's Manual of Systematic Bacteriology. PHA S, S M, ME S, JG H, editors. Baltimore: Williams & Wilkins; 1986.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
