

Review
doi: 10.1038/nrc2675.
Epub 2009 Jul 16.
Crosstalk of Notch with p53 and p63 in cancer growth control
Affiliations
- PMID: 19609265
- PMCID: PMC6059364
- DOI: 10.1038/nrc2675
Item in Clipboard
Review
Crosstalk of Notch with p53 and p63 in cancer growth control
Nat Rev Cancer.
2009 Aug.
Abstract
Understanding the complexity of cancer depends on an elucidation of the underlying regulatory networks, at the cellular and intercellular levels and in their temporal dimension. This Opinion article focuses on the multilevel crosstalk between the Notch pathway and the p53 and p63 pathways. These two coordinated signalling modules are at the interface of external damaging signals and control of stem cell potential and differentiation. Positive or negative reciprocal regulation of the two pathways can vary with cell type and cancer stage. Therefore, selective or combined targeting of the two pathways could improve the efficacy and reduce the toxicity of cancer therapies.
Figures
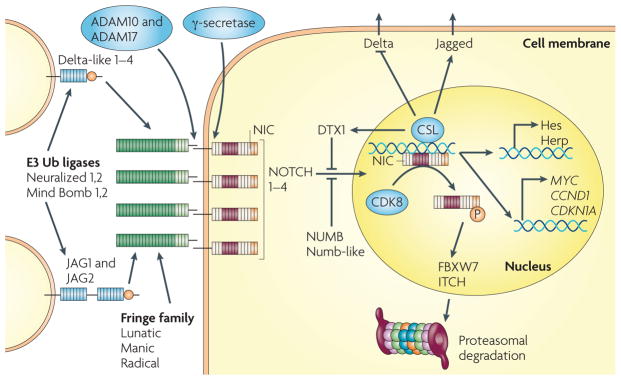
Two families of transmembrane Notch ligands and four Notch receptors exist in mammalian cells. The ligands must be ubiquitylated by specific e3 ubiquitin (Ub) ligases to induce Notch activity, which they have been proposed to do through several mechanisms, such as exerting a pulling force on the engaged receptor, ligand clustering or internalizing, and undergoing modification and subsequent reinsertion into the membrane. Modification of the extracellular domain of Notch receptors by Fringe family glycosyltransferases allows differential activation by the Delta or Jagged ligands. Following ligand engagement, Notch receptors are cleaved first by an ADAM metalloproteinase and then, at the juxta-membrane region, by the presenilin–γ-secretase complex. The resulting Notch intracellular domain (Nic) translocates to the nucleus, where it associates with the DNA binding protein csL (also known as rBPJκ), converting it from a repressor to an activator of transcription. This induces the transcription of two classes of target genes. Genes of the Hes family, which encode basic helix–loop–helix transcriptional repressors, and genes of the Herp family, which encode related but distinct basic helix–loop–helix proteins that heterodimerize and cooperate with Hes family members, are ubiquitously induced. cell cycle regulatory genes, such as MYC, CCND1 (which encodes cyclin D1) and CDKN1A (which encodes p21) are induced in a cell type-specific manner. in parallel, Notch activation controls neighbouring cells, through an opposite modulation of ligands of the Delta and Jagged families, which can provide a basis for lateral inhibition or lateral induction, respectively (as described in the main text). The Notch transcription-activating complex is terminated by phosphorylation of activated nuclear Notch by cyclin-dependent kinase 8 (cDK8), with subsequent recognition by the ubiquitin ligase FBXW7 (also known as seL-10) and proteasome-dependent degradation. Negative regulatory mechanisms are also provided by the e3 ubiquitin ligases Deltex 1 (DTX1) and iTcH and by NUMB, an asymmetrically distributed protein that has been linked to the recruitment of Notch into endocytic vescicles but also to NOTcH ubiquitylation. For more detailed information, see REFS ,,.
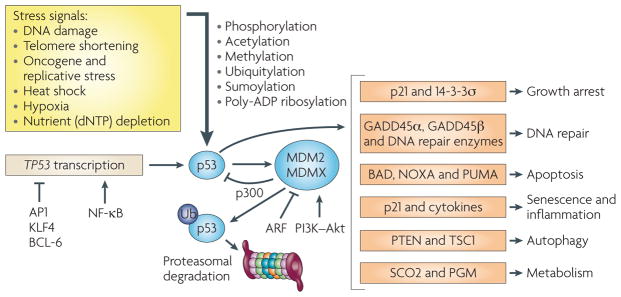
p53 activity is induced by many stress-related signals and is under the control of various transcriptional and post-transcriptional regulatory mechanisms. These include acetylation, methylation, phosphorylation and poly-ADP ribosylation, which have been implicated in the control of p53 nuclear versus cytoplasmic localization, as well as gene target selectivity. Phosphorylation of p53 at specific serine and threonine residues affects recognition by the ubiquitin (Ub) e3 ligase MDM2 and the related protein MDMX (also known as MDM4). Although the main function of MDM2 is to decrease p53 stability, MDM2 and MDMX can also suppress p53 transcriptional activity by competing with the acetyltransferase p300 for p53 binding. The role of p53 phosphorylation in vivo remains to be established, but the negative regulatory function of MDM2 and MDMX has been confirmed by mouse genetic models and by the use of small molecules, such as Nutlin-3, that block p53–MDM2 interactions. As MDM2 is one of the direct transcriptional targets of p53, the short and long arms of the negative feedback loop resulting from p53-dependent induction of MDM2 and MDM2-dependent suppression of p53 activity provide the basis for the oscillatory behaviour of the p53 pathway in response to DNA damage. For more detailed information on the p53 pathway, as well as the consequences of TP53 mutations not covered in this article, see REFS –,. AP1, activator protein 1; BAD, BcL-2-associated agonist of cell death; GADD45a, growth arrest and DNA-damage-inducible-a; KLF4, Kruppel-like factor 4; NF-κB, nuclear factor-κB; PGM, phosphoglycerate mutase; scO2, synthesis of cytochrome c oxidase 2; Tsc1, tuberous sclerosis 1.
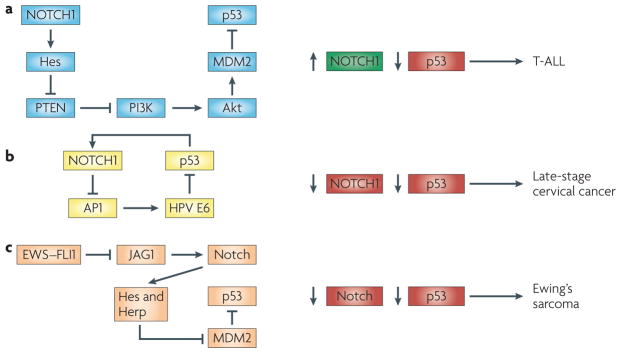
a | The oncogenic function of Notch signalling in T cell acute lymphoblastic leukeamia (T-ALL) is connected with the downmodulation of p53 through increased MDM2 activity. As discussed in the main text, a proposed mechanism, based on experimental and clinical studies–, involves activation of the Pi3K–Akt pathway through Hes family-mediated down-modulation of PTeN expression. b | A positive feedback loop between p53 and NOTcH1 can counteract cervical carcinogenesis. Advanced stages of cervical carcinogenesis are linked to a concomitant upregulation of the human papillomavirus (HPv) e6 oncoprotein and downmodulation of NOTCH1. p53 protein levels and activity are increased by NOTcH1 activation in cervical carcinoma cells through the suppression of activator protein 1 (AP1)-dependent transcription of HPv e6. in turn, NOTCH1 is a direct positive target and is induced by increased p53. c | in ewing’s sarcoma, the EWSR1–friend leukaemia virus integration 1 (FLI1) translocation product (eWs–FLi1) is a negative regulator of Jagged1 (JAG1) expression, with consequent suppression of Notch activity. The Notch pathway in this system positively regulates p53 activity through a mechanism involving transcriptional repression of MDM2 by Hes and Herp family transcriptional repressors.
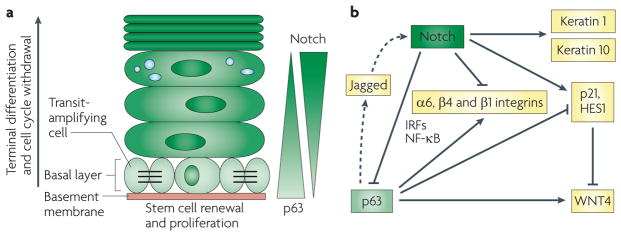
reciprocal feedback and feedforward loops between Notch and p63 contribute to the balance between self-renewing keratinocyte populations and cells at various stages of commitment to differentiation. a | in the interfollicular epidermis, keratinocytes undergo a vertical programme of differentiation, as they proceed from the basal proliferating compartment to the upper terminally differentiating layers. in the proliferative compartment, a dynamic equilibrium probably exists between a minor fraction of slow cycling cells with increased self-renewal potential (putative stem cells) and already-committed populations that are actively dividing but only for a limited number of times (transit-amplifying cells). Opposing gradients of p63 expression and Notch activity exist in the basal but not the upper epidermal layers. This may result, in part, from their reciprocal negative regulation. b | Mechanistically, increased Notch activity suppresses the expression of p63 through modulation of interferon-responsive factors (irFs) and nuclear factor-κB (NF-κB) signalling and, possibly, microrNAs. in turn, p63 binds to the HES1, CDKN1A (which encodes p21) and WNT4 promoters, counteracting the Notch effects on their expression (WNT4 is suppressed by Notch activation through an indirect mechanism mediated by both Hes1 and p21). p63 and Notch also increase and suppress, respectively, integrin expression (a6, β4 and β1 integrins), through less well-defined mechanisms. in parallel with these antagonistic effects, p63 can also synergize with Notch signalling in the early steps of differentiation (characterized by keratin 1 and keratin 10 expression), through a paracrine mechanism (illustrated by dotted arrows) involving the induction of expression of Jagged ligands and Notch activation in neighbouring cells. Downmodulation of p63 by increased Notch signalling could then be a signal for later differentiation stages to occur.
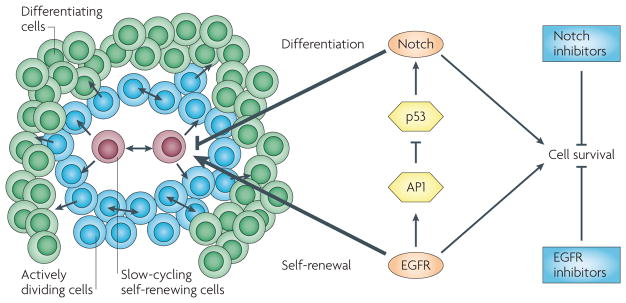
As in normal keratinocytes, in skin squamous cell carcinomas (illustrated on the left of the figure as an agglomerate of cells at various stages of the differentiation process) epidermal growth factor receptor (eGFr) signalling downmodulates NOTcH1 expression through activator protein 1 (AP1)-dependent suppression of TP53 transcription. squamous cell carcinomas are intrinsically heterogeneous and contain cells that have increased self-renewing potential but are possibly slow-cycling, actively dividing cells and cells that undergo differentiation. eGFr promotes self-renewal by increasing the proliferation or survival of both slow-cycling and actively dividing cells. conversely, Notch restricts proliferation potential and promotes differentiation. Although eGFr and Notch can exert opposite effects on differentiation, they both increase cell survival. As a result, pharmacological inhibition of both pathways can synergize in the induction of apoptosis.
References
-
- Stelling J, Sauer U, Szallasi Z, Doyle FJ, Doyle J. Robustness of cellular functions. Cell. 2004;118:675–685. - PubMed
-
- Alon U. Network motifs: theory and experimental approaches. Nature Rev Genet. 2007;8:450–461. - PubMed
-
- Bray SJ. Notch signalling: a simple pathway becomes complex. Nature Rev Mol Cell Biol. 2006;7:678–689. - PubMed
-
- Hurlbut GD, Kankel MW, Lake RJ, Artavanis-Tsakonas S. Crossing paths with Notch in the hyper-network. Curr Opin Cell Biol. 2007;19:166–175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
