

Evolutionary dynamics of human Toll-like receptors and their different contributions to host defense
- PMID: 19609346
- PMCID: PMC2702086
- DOI: 10.1371/journal.pgen.1000562
Evolutionary dynamics of human Toll-like receptors and their different contributions to host defense
Abstract
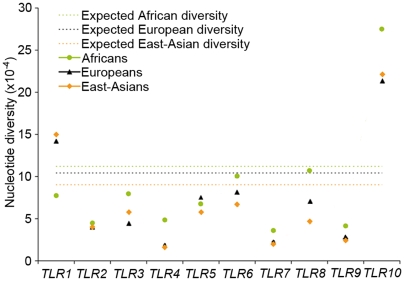
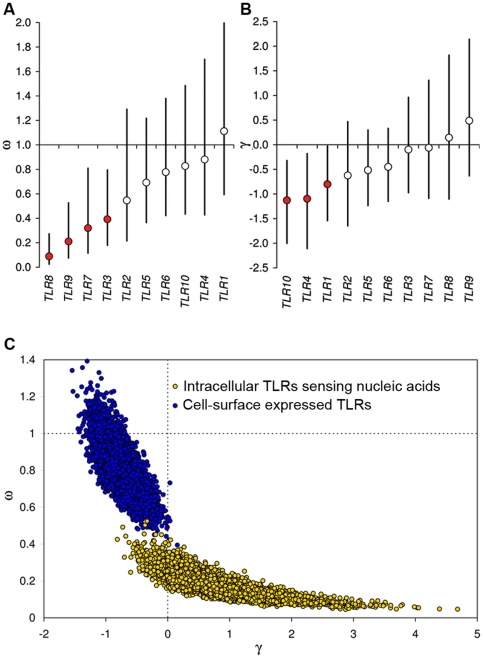
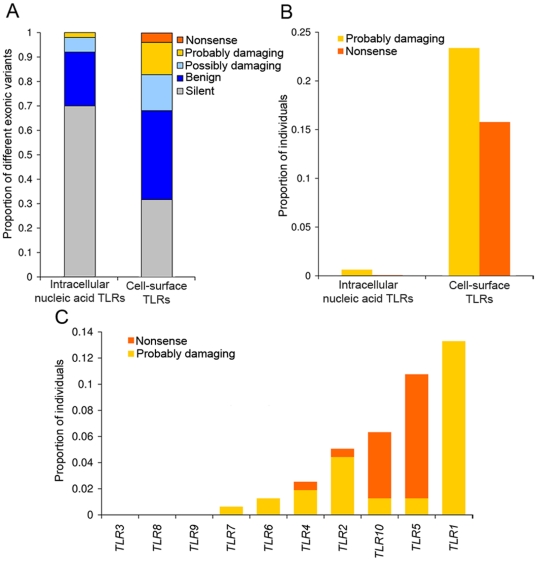
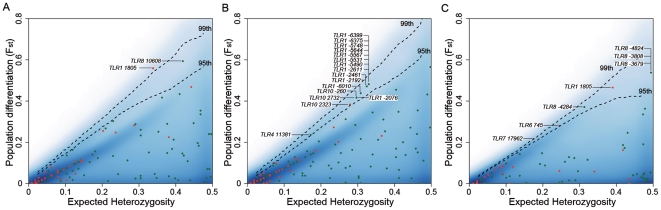
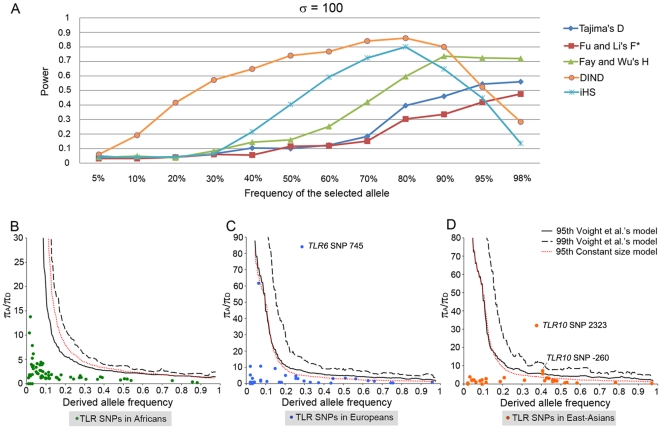
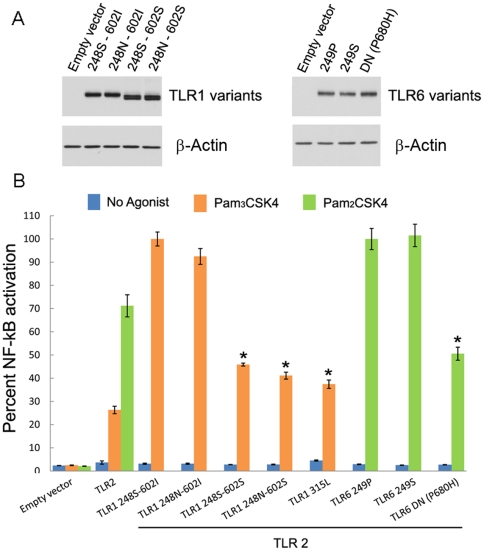
Infectious diseases have been paramount among the threats to health and survival throughout human evolutionary history. Natural selection is therefore expected to act strongly on host defense genes, particularly on innate immunity genes whose products mediate the direct interaction between the host and the microbial environment. In insects and mammals, the Toll-like receptors (TLRs) appear to play a major role in initiating innate immune responses against microbes. In humans, however, it has been speculated that the set of TLRs could be redundant for protective immunity. We investigated how natural selection has acted upon human TLRs, as an approach to assess their level of biological redundancy. We sequenced the ten human TLRs in a panel of 158 individuals from various populations worldwide and found that the intracellular TLRs -- activated by nucleic acids and particularly specialized in viral recognition -- have evolved under strong purifying selection, indicating their essential non-redundant role in host survival. Conversely, the selective constraints on the TLRs expressed on the cell surface -- activated by compounds other than nucleic acids -- have been much more relaxed, with higher rates of damaging nonsynonymous and stop mutations tolerated, suggesting their higher redundancy. Finally, we tested whether TLRs have experienced spatially-varying selection in human populations and found that the region encompassing TLR10-TLR1-TLR6 has been the target of recent positive selection among non-Africans. Our findings indicate that the different TLRs differ in their immunological redundancy, reflecting their distinct contributions to host defense. The insights gained in this study foster new hypotheses to be tested in clinical and epidemiological genetics of infectious disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Lemaitre B, Hoffmann J. The host defense of Drosophila melanogaster. Annu Rev Immunol. 2007;25:697–743. - PubMed
-
- Janeway CA, Jr., Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197–216. - PubMed
-
- Medzhitov R, Janeway CA., Jr. Innate immunity: the virtues of a nonclonal system of recognition. Cell. 1997;91:295–298. - PubMed
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Beutler B, Rietschel ET. Innate immune sensing and its roots: the story of endotoxin. Nat Rev Immunol. 2003;3:169–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
