

Two chromatin remodeling activities cooperate during activation of hormone responsive promoters
- PMID: 19609353
- PMCID: PMC2704372
- DOI: 10.1371/journal.pgen.1000567
Two chromatin remodeling activities cooperate during activation of hormone responsive promoters
Abstract
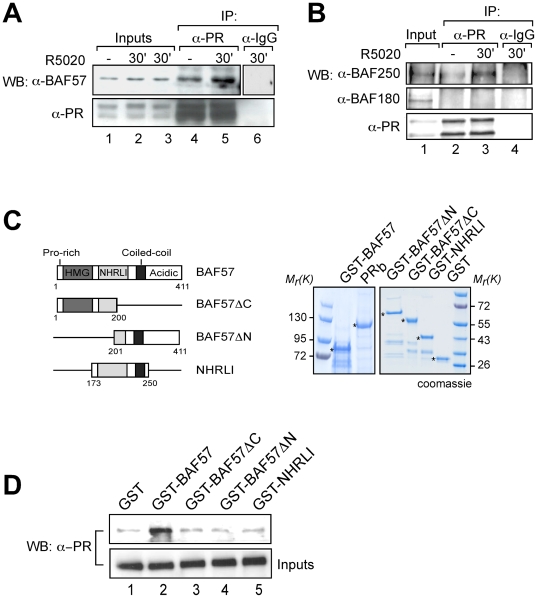
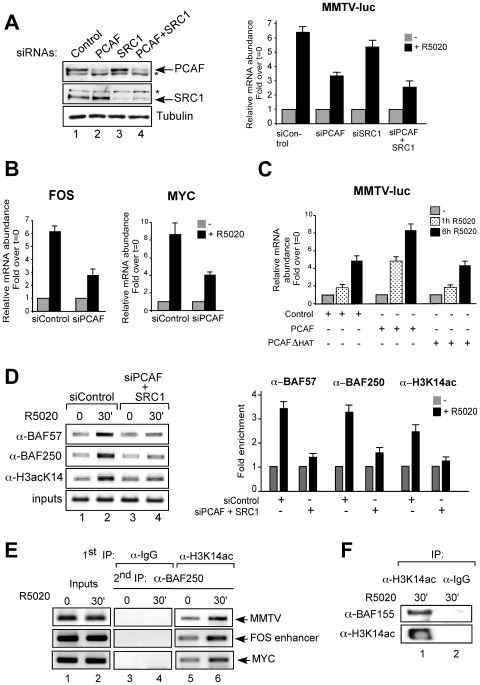
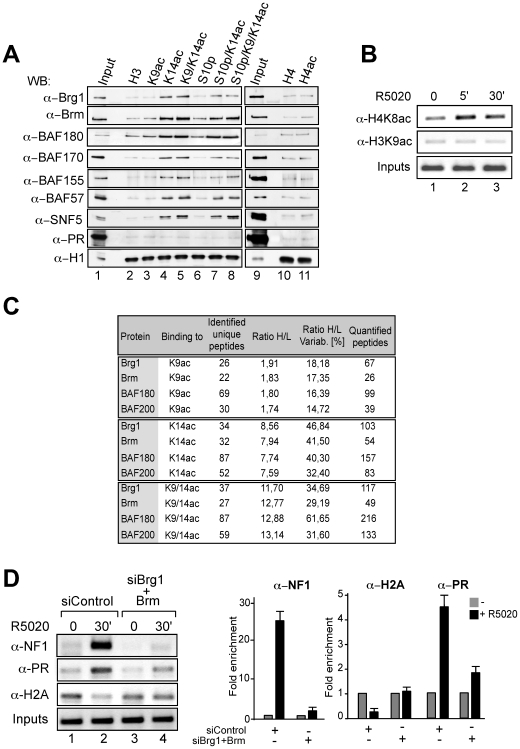
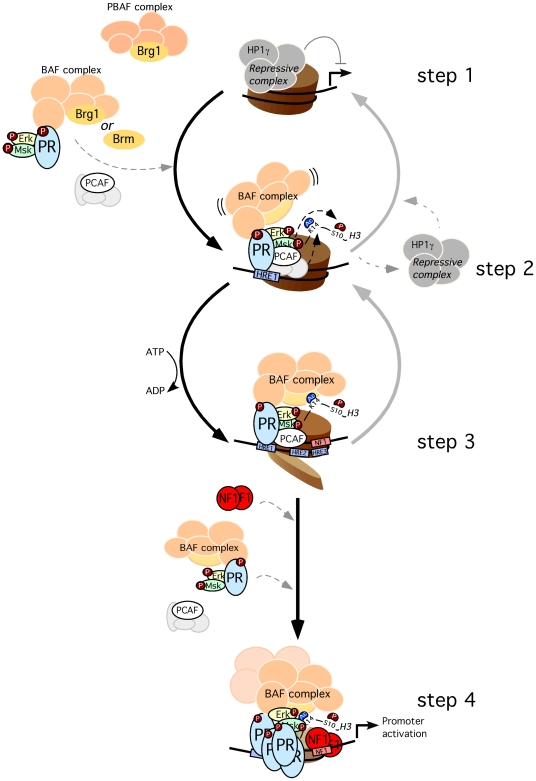
Steroid hormones regulate gene expression by interaction of their receptors with hormone responsive elements (HREs) and recruitment of kinases, chromatin remodeling complexes, and coregulators to their target promoters. Here we show that in breast cancer cells the BAF, but not the closely related PBAF complex, is required for progesterone induction of several target genes including MMTV, where it catalyzes localized displacement of histones H2A and H2B and subsequent NF1 binding. PCAF is also needed for induction of progesterone target genes and acetylates histone H3 at K14, an epigenetic mark that interacts with the BAF subunits by anchoring the complex to chromatin. In the absence of PCAF, full loading of target promoters with hormone receptors and BAF is precluded, and induction is compromised. Thus, activation of hormone-responsive promoters requires cooperation of at least two chromatin remodeling activities, BAF and PCAF.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
-
- Aalfs JD, Kingston RE. What does ‘chromatin remodeling’ mean? Trends Biochem Sci. 2000;25:548–555. - PubMed
-
- Whitehouse I, Rando OJ, Delrow J, Tsukiyama T. Chromatin remodelling at promoters suppresses antisense transcription. Nature. 2007;450:1031–1035. - PubMed
-
- Mohrmann L, Verrijzer CP. Composition and functional specificity of SWI2/SNF2 class chromatin remodeling complexes. Biochim Biophys Acta. 2005;1681:59–73. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
