

Conformational dynamics in the Acyl-CoA synthetases, adenylation domains of non-ribosomal peptide synthetases, and firefly luciferase
- PMID: 19610673
- PMCID: PMC2769252
- DOI: 10.1021/cb900156h
Conformational dynamics in the Acyl-CoA synthetases, adenylation domains of non-ribosomal peptide synthetases, and firefly luciferase
Abstract
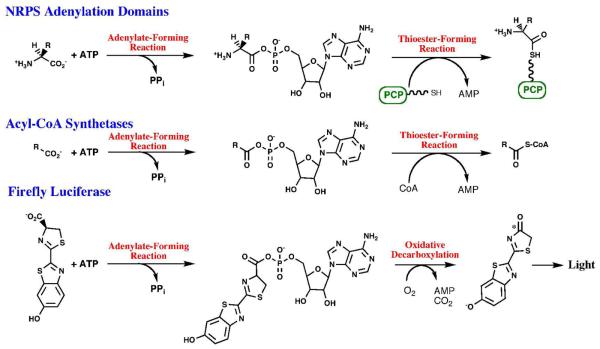
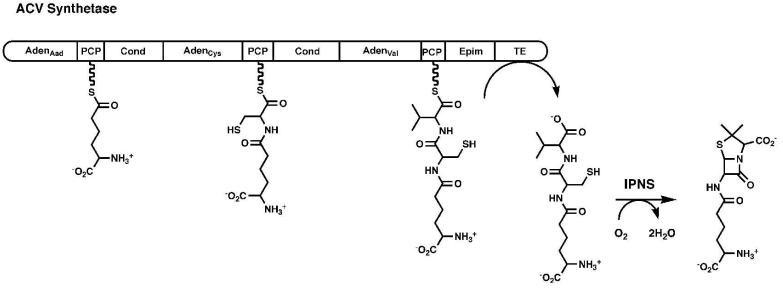
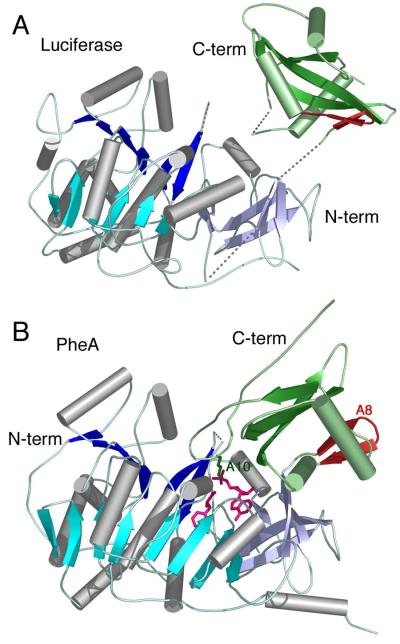
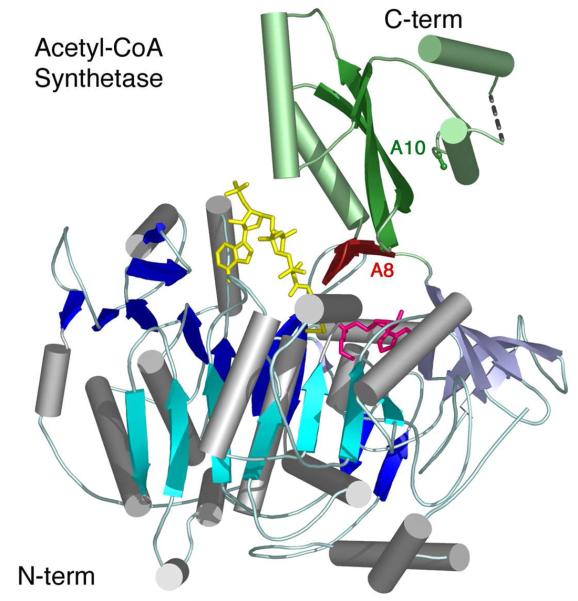
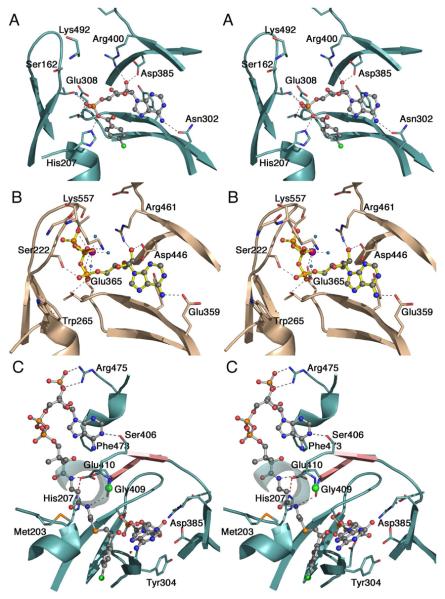
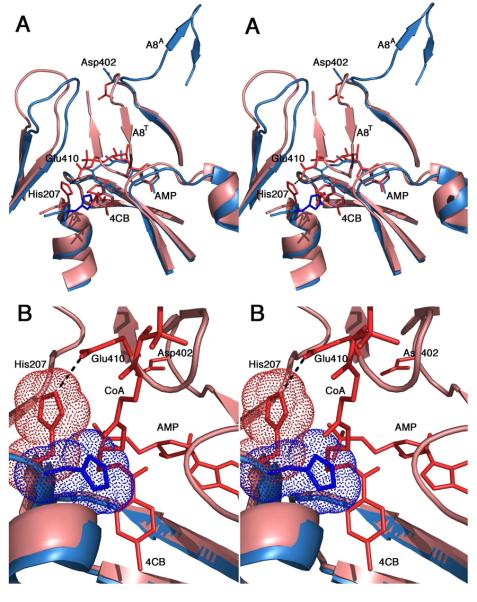
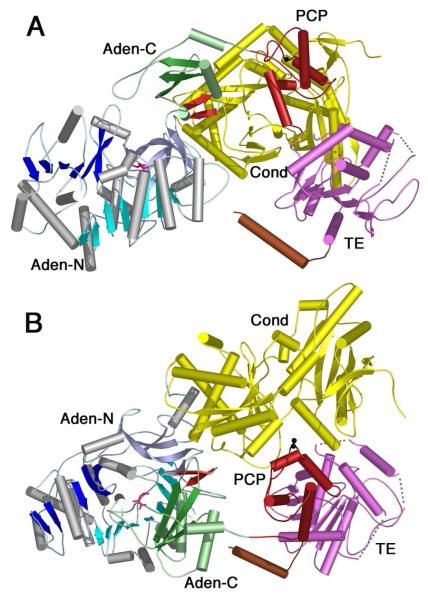
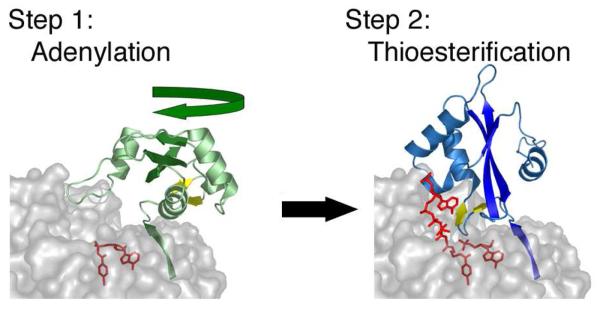
The ANL superfamily of adenylating enzymes contains acyl- and aryl-CoA synthetases, firefly luciferase, and the adenylation domains of the modular non-ribosomal peptide synthetases (NRPSs). Members of this family catalyze two partial reactions: the initial adenylation of a carboxylate to form an acyl-AMP intermediate, followed by a second partial reaction, most commonly the formation of a thioester. Recent biochemical and structural evidence has been presented that supports the use by this enzyme family of a remarkable catalytic strategy for the two catalytic steps. The enzymes use a 140 degrees domain rotation to present opposing faces of the dynamic C-terminal domain to the active site for the different partial reactions. Support for this domain alternation strategy is presented along with an explanation of the advantage of this catalytic strategy for the reaction catalyzed by the ANL enzymes. Finally, the ramifications of this domain rotation in the catalytic cycle of the modular NRPS enzymes are discussed.
Figures
References
-
- McElroy WD, DeLuca M, Travis J. Molecular uniformity in biological catalyses. The enzymes concerned with firefly luciferin, amino acid, and fatty acid utilization are compared. Science. 1967;157:150–160. - PubMed
-
- Lipmann F. Enzymatic synthesis of acetyl phosphate. J Biol Chem. 1944;155:55–70.
-
- Berg P. Acyl adenylates; an enzymatic mechanism of acetate activation. J Biol Chem. 1956;222:991–1013. - PubMed
-
- Farrar WW, Plowman KM. Kinetics of acetyl-CA synthetase–1. Mode of addition of substrates. International J. Biochem. 1975;6:537–542.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
