

Corticostriatal dysfunction underlies diminished striatal ascorbate release in the R6/2 mouse model of Huntington's disease
- PMID: 19616518
- PMCID: PMC2745264
- DOI: 10.1016/j.brainres.2009.07.019
Corticostriatal dysfunction underlies diminished striatal ascorbate release in the R6/2 mouse model of Huntington's disease
Abstract
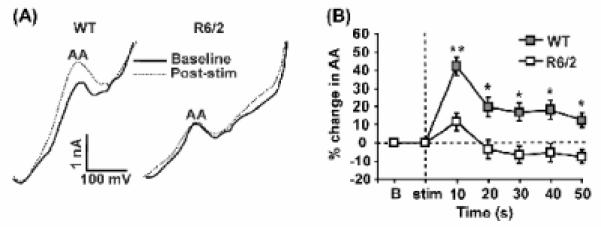
A behavior-related deficit in the release of ascorbate (AA), an antioxidant vitamin, occurs in the striatum of R6/2 mice expressing the human mutation for Huntington's disease (HD), a dominantly inherited condition characterized by striatal dysfunction. To determine the role of corticostriatal fibers in AA release, we combined slow-scan voltammetry with electrical stimulation of cortical afferents to measure evoked fluctuations in extracellular AA in wild-type (WT) and R6/2 striatum. Although cortical stimulation evoked a rapid increase in AA release in both groups, the R6/2 response had a significantly shorter duration and smaller magnitude than WT. To determine if corticostriatal dysfunction also underlies the behavior-related AA deficit in R6/2s, we measured striatal AA release in separate groups of mice treated with d-amphetamine (5 mg/kg), a psychomotor stimulant known to release AA from corticostriatal terminals independently of dopamine. Relative to WT, both AA release and behavioral activation were diminished in R6/2 mice. Collectively, our results show that the corticostriatal pathway is directly involved in AA release and that this system is dysfunctional in HD. Moreover, because AA release requires glutamate uptake, a failure of striatal AA release in HD is consistent with an overactive glutamate system and diminished glutamate transport, both of which are thought to be central to HD pathogenesis.
Figures


Similar articles
-
Up-regulation of GLT1 reverses the deficit in cortically evoked striatal ascorbate efflux in the R6/2 mouse model of Huntington's disease.J Neurochem. 2012 May;121(4):629-38. doi: 10.1111/j.1471-4159.2012.07691.x. Epub 2012 Mar 28. J Neurochem. 2012. PMID: 22332910 Free PMC article.
-
Dysregulation of corticostriatal ascorbate release and glutamate uptake in transgenic models of Huntington's disease.Antioxid Redox Signal. 2013 Dec 10;19(17):2115-28. doi: 10.1089/ars.2013.5387. Epub 2013 Jun 19. Antioxid Redox Signal. 2013. PMID: 23642110 Free PMC article.
-
Dysregulation of ascorbate release in the striatum of behaving mice expressing the Huntington's disease gene.J Neurosci. 2002 Jan 15;22(2):RC202. doi: 10.1523/JNEUROSCI.22-02-j0006.2002. J Neurosci. 2002. PMID: 11784814 Free PMC article.
-
Corticostriatal network dysfunction in Huntington's disease: Deficits in neural processing, glutamate transport, and ascorbate release.CNS Neurosci Ther. 2018 Apr;24(4):281-291. doi: 10.1111/cns.12828. Epub 2018 Feb 21. CNS Neurosci Ther. 2018. PMID: 29464896 Free PMC article. Review.
-
Dysregulation of Corticostriatal Connectivity in Huntington's Disease: A Role for Dopamine Modulation.J Huntingtons Dis. 2016 Dec 15;5(4):303-331. doi: 10.3233/JHD-160221. J Huntingtons Dis. 2016. PMID: 27983564 Free PMC article. Review.
Cited by
-
The Role of Vitamins in Neurodegenerative Disease: An Update.Biomedicines. 2021 Sep 22;9(10):1284. doi: 10.3390/biomedicines9101284. Biomedicines. 2021. PMID: 34680401 Free PMC article. Review.
-
Up-regulation of GLT1 reverses the deficit in cortically evoked striatal ascorbate efflux in the R6/2 mouse model of Huntington's disease.J Neurochem. 2012 May;121(4):629-38. doi: 10.1111/j.1471-4159.2012.07691.x. Epub 2012 Mar 28. J Neurochem. 2012. PMID: 22332910 Free PMC article.
-
Old Things New View: Ascorbic Acid Protects the Brain in Neurodegenerative Disorders.Int J Mol Sci. 2015 Nov 27;16(12):28194-217. doi: 10.3390/ijms161226095. Int J Mol Sci. 2015. PMID: 26633354 Free PMC article. Review.
-
BDNF-TrkB signaling in striatopallidal neurons controls inhibition of locomotor behavior.Nat Commun. 2013;4:2031. doi: 10.1038/ncomms3031. Nat Commun. 2013. PMID: 23774276 Free PMC article.
-
Characterization of neurophysiological and behavioral changes, MRI brain volumetry and 1H MRS in zQ175 knock-in mouse model of Huntington's disease.PLoS One. 2012;7(12):e50717. doi: 10.1371/journal.pone.0050717. Epub 2012 Dec 20. PLoS One. 2012. PMID: 23284644 Free PMC article.
References
-
- Ariano MA, Aronin N, Difiglia M, Tagle DA, Sibley DR, Leavitt BR, Hayden MR, Levine MS. Striatal neurochemical changes in transgenic models of Huntington’s disease. J Neurosci Res. 2002;68:716–29. - PubMed
-
- Basse-Tomusk A, Rebec GV. Corticostriatal and thalamic regulation of amphetamine-induced ascorbate release in the neostriatum. Pharmacol Biochem Behav. 1990;35:55–60. - PubMed
-
- Behrens PF, Franz P, Woodman B, Lindenberg KS, Landwehrmeyer GB. Impaired glutamate transport and glutamate-glutamine cycling: downstream effects of the Huntington mutation. Brain. 2002;125:1908–22. - PubMed
-
- Berger UV, Hediger MA. The vitamin C transporter is expressed by astrocytes in culture but not in situ. 2000;11:1395–9. - PubMed
-
- Browne SE, Beal MF. Oxidative damage in Huntington’s disease pathogenesis. Antioxid Redox Signal. 2006;8:2061–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
