

Cell fate decision mediated by p53 pulses
- PMID: 19617533
- PMCID: PMC2710988
- DOI: 10.1073/pnas.0813088106
Cell fate decision mediated by p53 pulses
Abstract
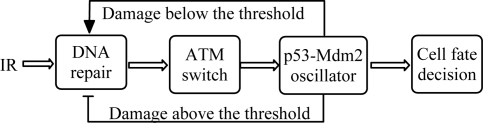
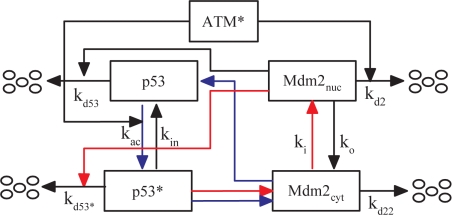
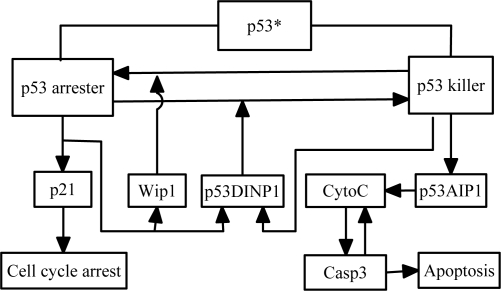
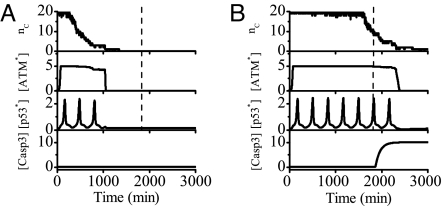
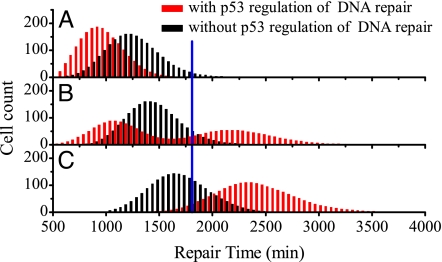
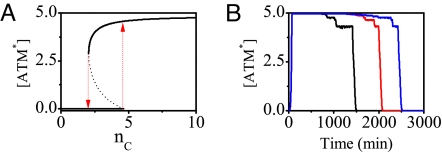
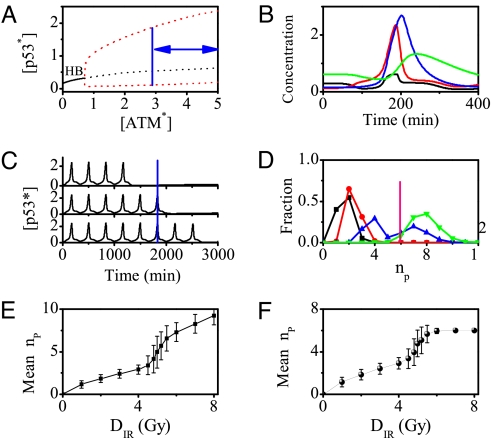
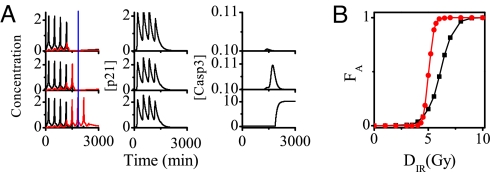
The tumor suppressor p53 plays a crucial role in cellular response to various stresses. Recent experiments have shown that p53 level exhibits a series of pulses after DNA damage caused by ionizing radiation (IR). However, how the p53 pulses govern cell survival and death remains unclear. Here, we develop an integrated model with four modules for the p53 network and explore the mechanism for cell fate decision based on the dynamics of the network. By numerical simulations, the following processes are characterized. First, DNA repair proteins bind to IR-induced double-strand breaks, forming complexes, which are then detected by ataxia telangiectasia mutated (ATM). Activated ATM initiates the p53 oscillator to produce pulses. Consequently, the target genes of p53 are selectively induced to control cell fate. We propose that p53 promotes the repair of minor DNA damage but suppresses the repair of severe damage. We demonstrate that cell fate is determined by the number of p53 pulses relying on the extent of DNA damage. At low damage levels, few p53 pulses evoke cell cycle arrest by inducing p21 and promote cell survival, whereas at high damage levels, sustained p53 pulses trigger apoptosis by inducing p53AIP1. We find that p53 can effectively maintain genomic integrity by regulating the efficiency and fidelity of DNA repair. We also show that stochasticity in the generation and repair of DNA damage leads to variability in cell fate. These findings are consistent with experimental observations and advance our understanding of the dynamics and functions of the p53 network.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Vousden KH, Lane DP. p53 in health and disease. Nat Rev Mol Cell Biol. 2007;8:275–283. - PubMed
-
- Harris SL, Levine AJ. The p53 pathway: positive and negative feedback loops. Oncogene. 2005;24:2899–2908. - PubMed
-
- Hu W, et al. A single nucleotide polymorphism in the Mdm2 gene disrupts the oscillation of p53 and Mdm2 levels in cells. Cancer Res. 2007;67:2757–2765. - PubMed
-
- Ramalingam S, et al. Quantitative assessment of the p53-Mdm2 feedback loop using protein lysate microarrays. Cancer Res. 2007;67:6247–6252. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
