

A hyperfused mitochondrial state achieved at G1-S regulates cyclin E buildup and entry into S phase
- PMID: 19617534
- PMCID: PMC2710990
- DOI: 10.1073/pnas.0904875106
A hyperfused mitochondrial state achieved at G1-S regulates cyclin E buildup and entry into S phase
Abstract
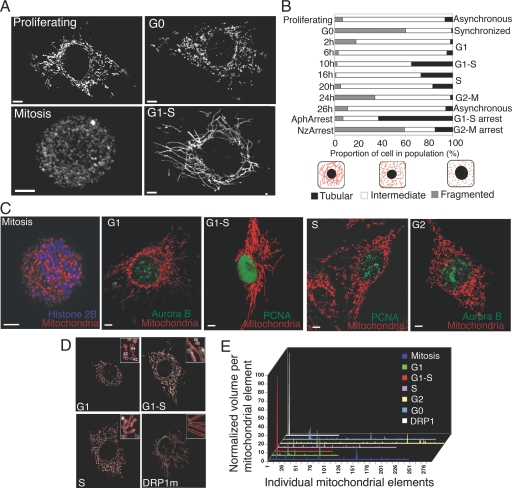
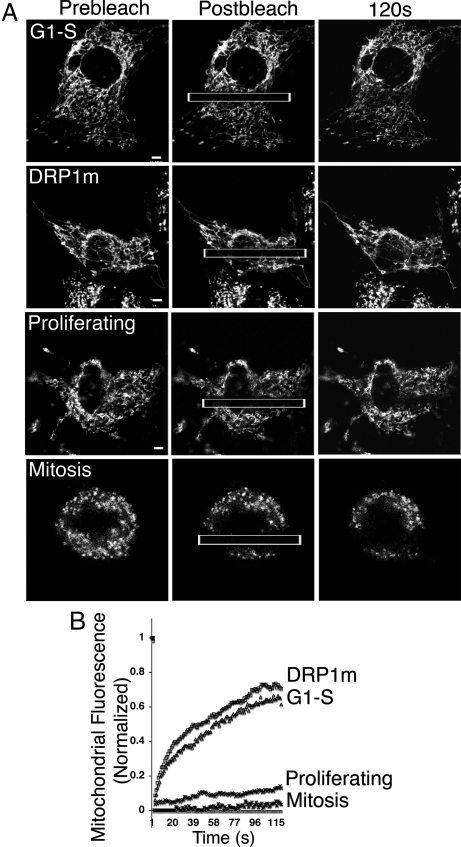
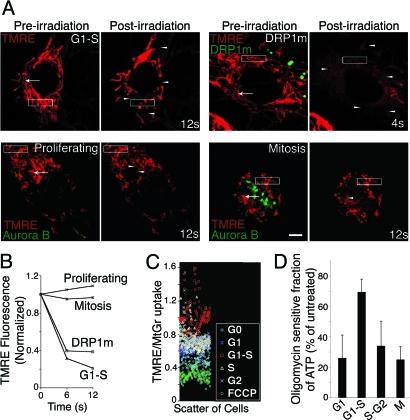
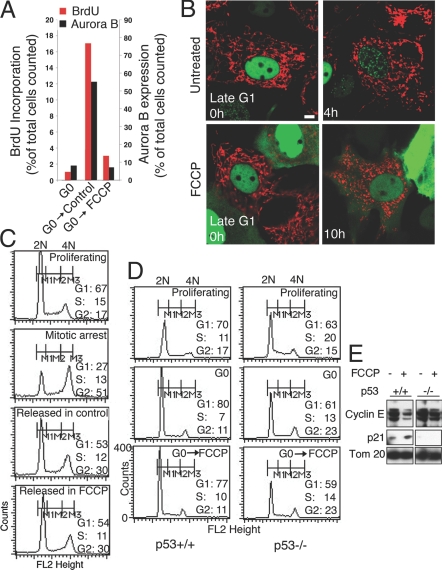
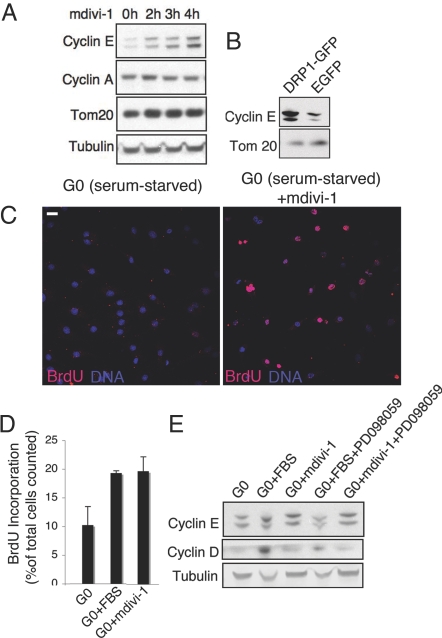
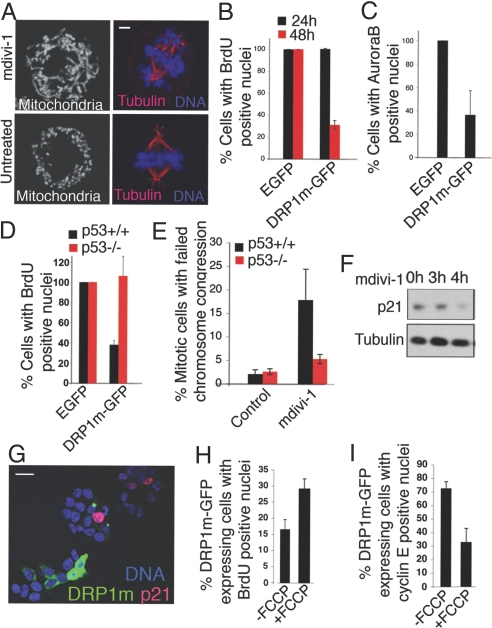
Mitochondria undergo fission-fusion events that render these organelles highly dynamic in cells. We report a relationship between mitochondrial form and cell cycle control at the G(1)-S boundary. Mitochondria convert from isolated, fragmented elements into a hyperfused, giant network at G(1)-S transition. The network is electrically continuous and has greater ATP output than mitochondria at any other cell cycle stage. Depolarizing mitochondria at early G(1) to prevent these changes causes cell cycle progression into S phase to be blocked. Inducing mitochondrial hyperfusion by acute inhibition of dynamin-related protein-1 (DRP1) causes quiescent cells maintained without growth factors to begin replicating their DNA and coincides with buildup of cyclin E, the cyclin responsible for G(1)-to-S phase progression. Prolonged or untimely formation of hyperfused mitochondria, through chronic inhibition of DRP1, causes defects in mitotic chromosome alignment and S-phase entry characteristic of cyclin E overexpression. These findings suggest a hyperfused mitochondrial system with specialized properties at G(1)-S is linked to cyclin E buildup for regulation of G(1)-to-S progression.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
The Krebs cycle meets the cell cycle: mitochondria and the G1-S transition.Proc Natl Acad Sci U S A. 2009 Jul 21;106(29):11825-6. doi: 10.1073/pnas.0906430106. Epub 2009 Jul 15. Proc Natl Acad Sci U S A. 2009. PMID: 19617546 Free PMC article. No abstract available.
References
-
- Detmer SA, Chan DC. Functions and dysfunctions of mitochondrial dynamics. Nat Rev Mol Cell Biol. 2007;8:870–879. - PubMed
-
- Karbowski M, Youle RJ. Dynamics of mitochondrial morphology in healthy cells and during apoptosis. Cell Death Differ. 2003;10:870–880. - PubMed
-
- Mandal S, Guptan P, Owusu-Ansah E, Banerjee U. Mitochondrial regulation of cell cycle progression during development as revealed by the tenured mutation in Drosophila. Dev Cell. 2005;9:843–854. - PubMed
-
- Matoba S, et al. p53 regulates mitochondrial respiration. Science. 2006;312:1650–1653. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
