

An examination of the relationship between active site loop size and thermodynamic activation parameters for orotidine 5'-monophosphate decarboxylase from mesophilic and thermophilic organisms
- PMID: 19618917
- PMCID: PMC2754381
- DOI: 10.1021/bi901064k
An examination of the relationship between active site loop size and thermodynamic activation parameters for orotidine 5'-monophosphate decarboxylase from mesophilic and thermophilic organisms
Abstract
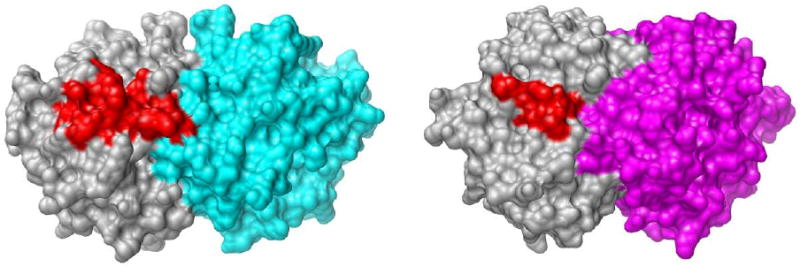
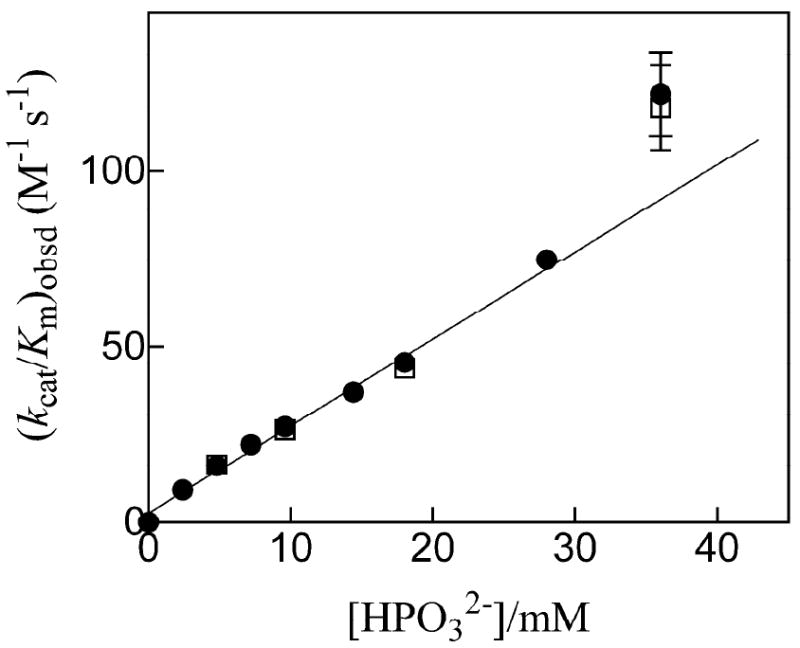
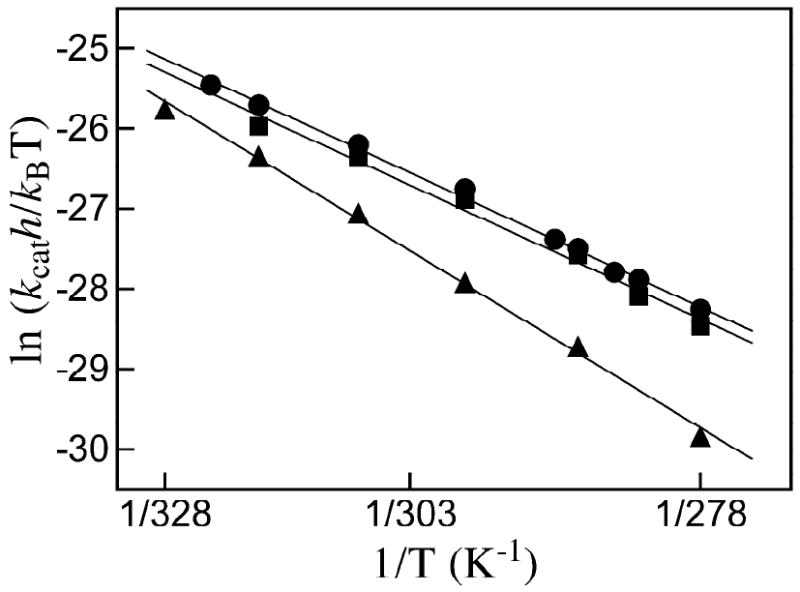
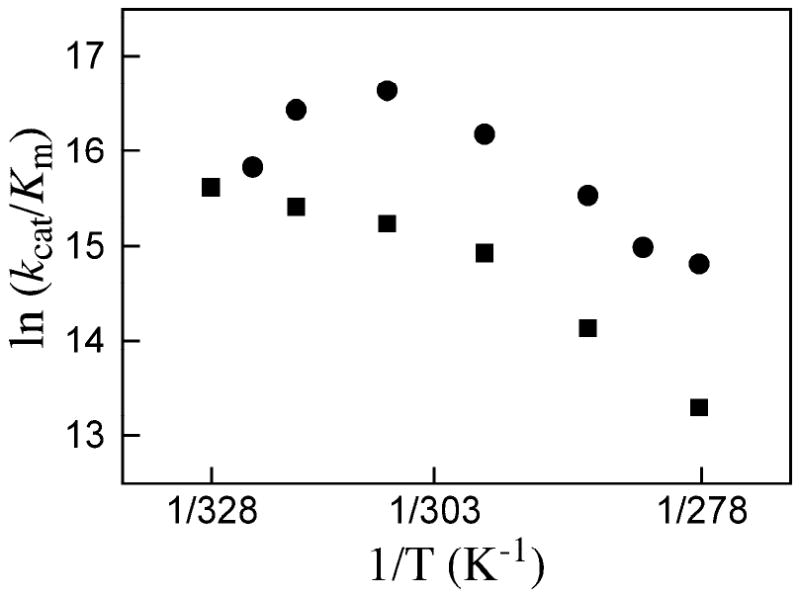
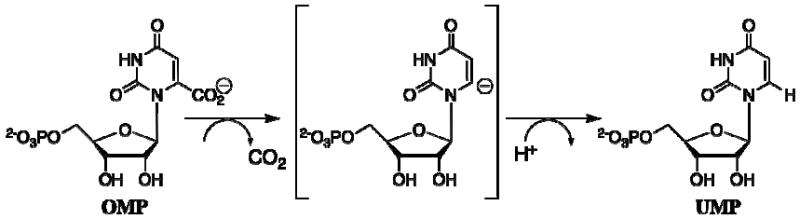
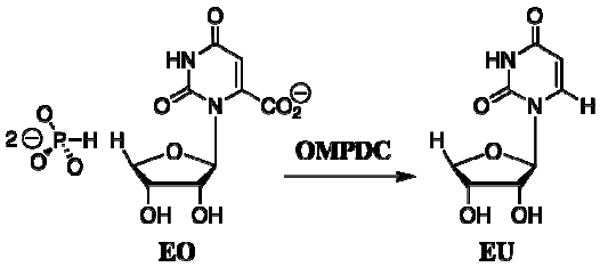
Closure of the active site phosphate gripper loop of orotidine 5'-monophosphate decarboxylase from Saccharomyces cerevisiae (ScOMPDC) over the bound substrate orotidine 5'-monophosphate (OMP) activates the bound substrate for decarboxylation by at least 10(4)-fold [Amyes, T. L., Richard, J. P., and Tait, J. J. (2005) J. Am. Chem. Soc. 127, 15708-15709]. The 19-residue phosphate gripper loop of the mesophilic ScOMPDC is much larger than the nine-residue loop at the ortholog from the thermophile Methanothermobacter thermautotrophicus (MtOMPDC). This difference in loop size results in a small decrease in the total intrinsic phosphate binding energy of the phosphodianion group of OMP from 11.9 to 11.6 kcal/mol, along with a modest decrease in the extent of activation by phosphite dianion of decarboxylation of the truncated substrate 1-(beta-D-erythrofuranosyl)orotic acid. The activation parameters DeltaH(double dagger) and DeltaS(double dagger) for k(cat) for decarboxylation of OMP are 3.6 kcal/mol and 10 cal K(-1) mol(-1) more positive, respectively, for MtOMPDC than for ScOMPDC. We suggest that these differences are related to the difference in the size of the active site loops at the mesophilic ScOMPDC and the thermophilic MtOMPDC. The greater enthalpic transition state stabilization available from the more extensive loop-substrate interactions for the ScOMPDC-catalyzed reaction is largely balanced by a larger entropic requirement for immobilization of the larger loop at this enzyme.
Figures
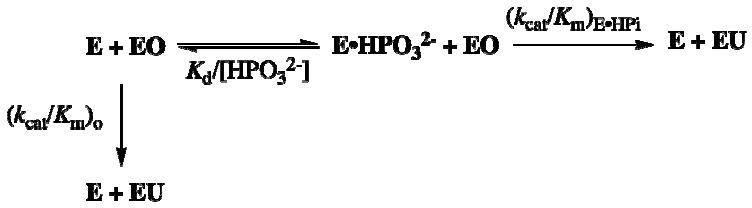
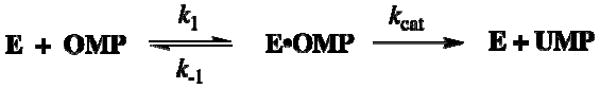
References
-
- Callahan BP, Miller BG. OMP decarboxylase - An enigma persists. Bioorg Chem. 2007;35:465–469. - PubMed
-
- Miller BG, Wolfenden R. Catalytic proficiency: the unusual case of OMP decarboxylase. Annu Rev Biochem. 2002;71:847–885. - PubMed
-
- Radzicka A, Wolfenden R. A proficient enzyme. Science. 1995;267:90–93. - PubMed
-
- Porter DJT, Short SA. Yeast Orotidine-5′-Phosphate Decarboxylase: Steady-State and Pre-Steady-State Analysis of the Kinetic Mechanism of Substrate Decarboxylation. Biochemistry. 2000;39:11788–11800. - PubMed
-
- Stanton CL, Kuo IF, Mundy CJ, Laino T, Houk KN. QM/MM metadynamics study of the direct decarboxylation mechanism for orotidine-5′-monophosphate decarboxylase using two different QM regions: acceleration too small to explain rate of enzyme catalysis. J Phys Chem B. 2007;111:12573–12581. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
