

The amino acid motif L/IIxxFE defines a novel actin-binding sequence in PDZ-RhoGEF
- PMID: 19618964
- PMCID: PMC2728795
- DOI: 10.1021/bi9010013
The amino acid motif L/IIxxFE defines a novel actin-binding sequence in PDZ-RhoGEF
Abstract
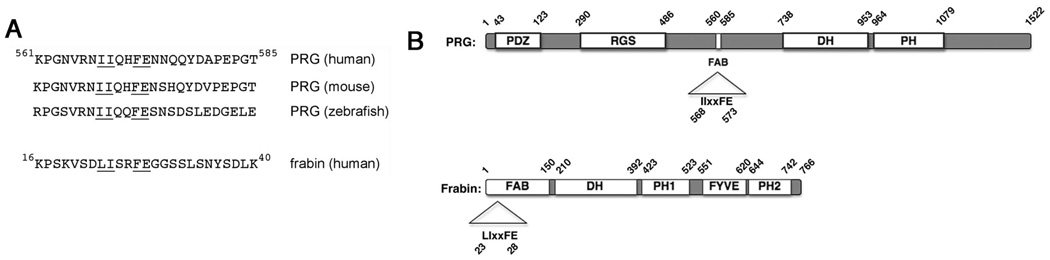
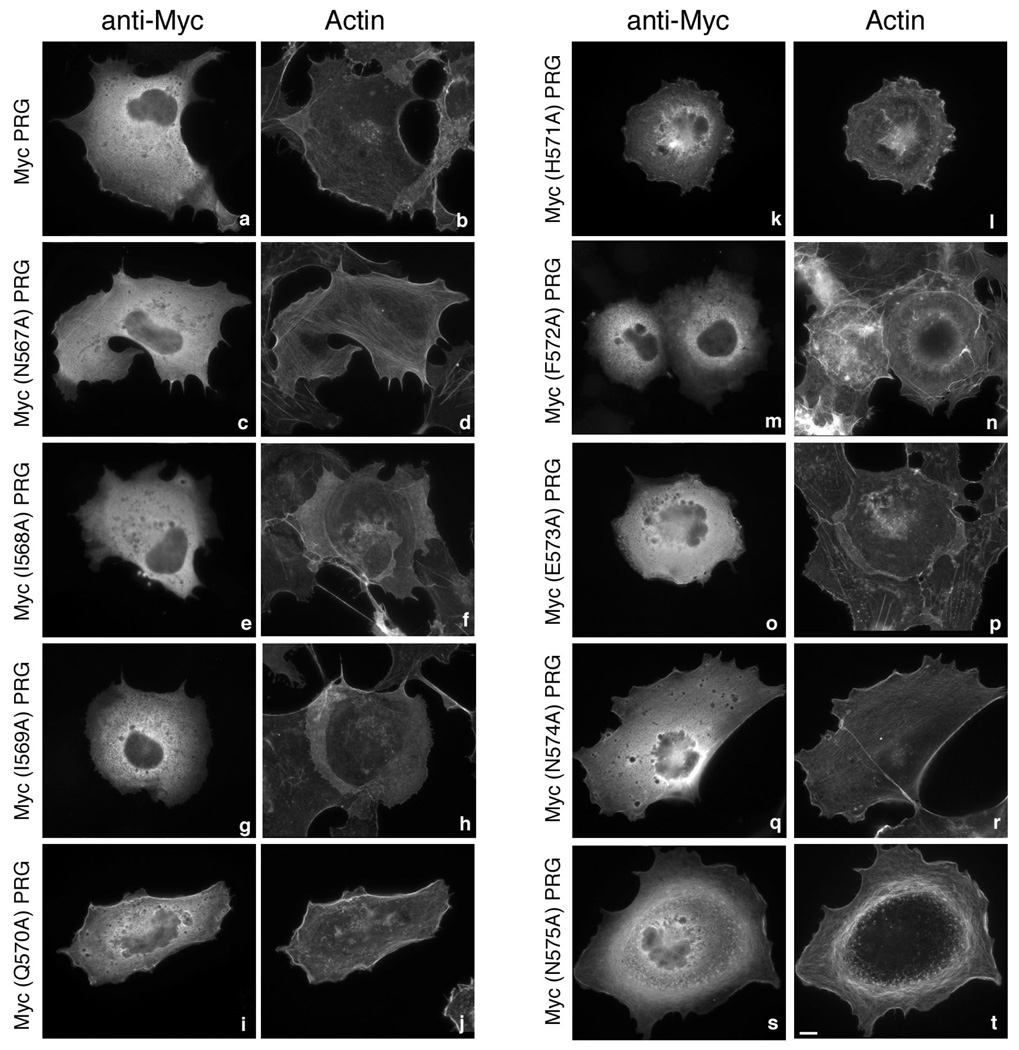
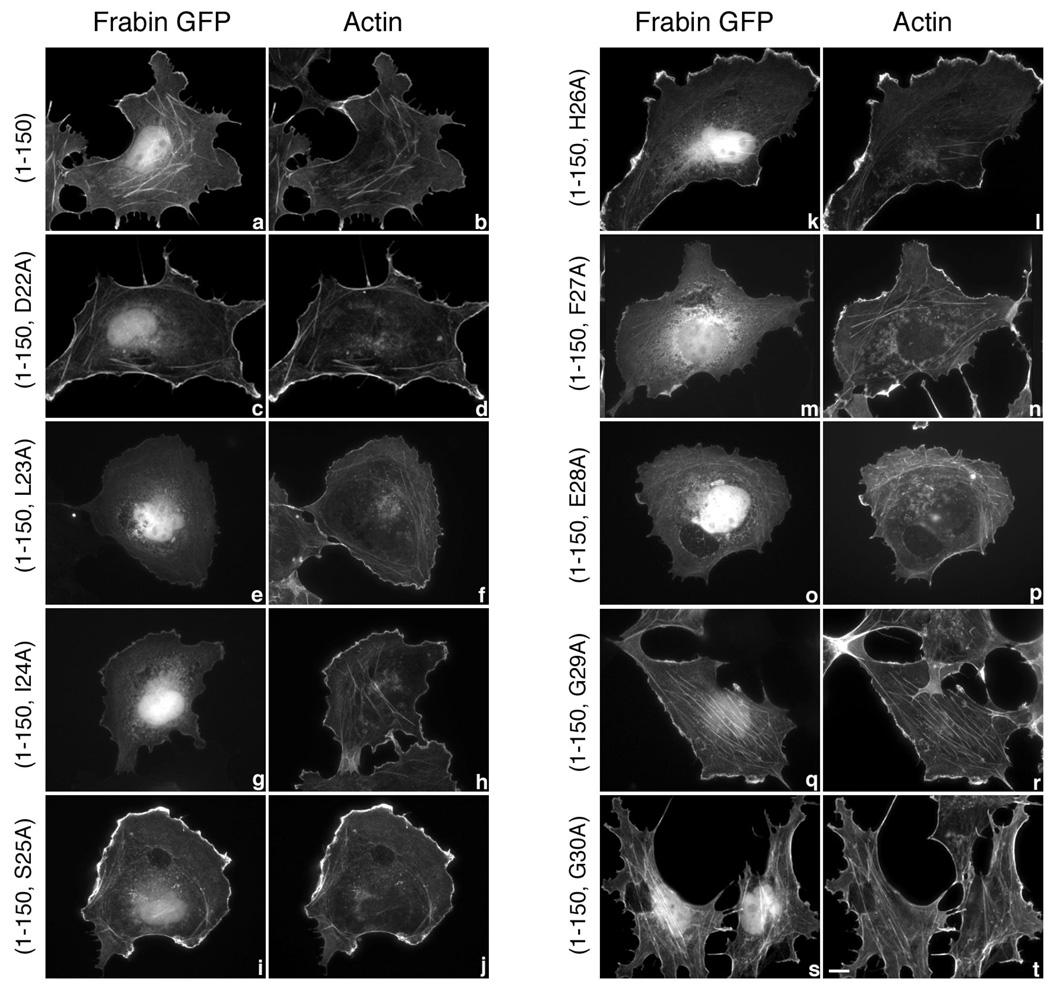
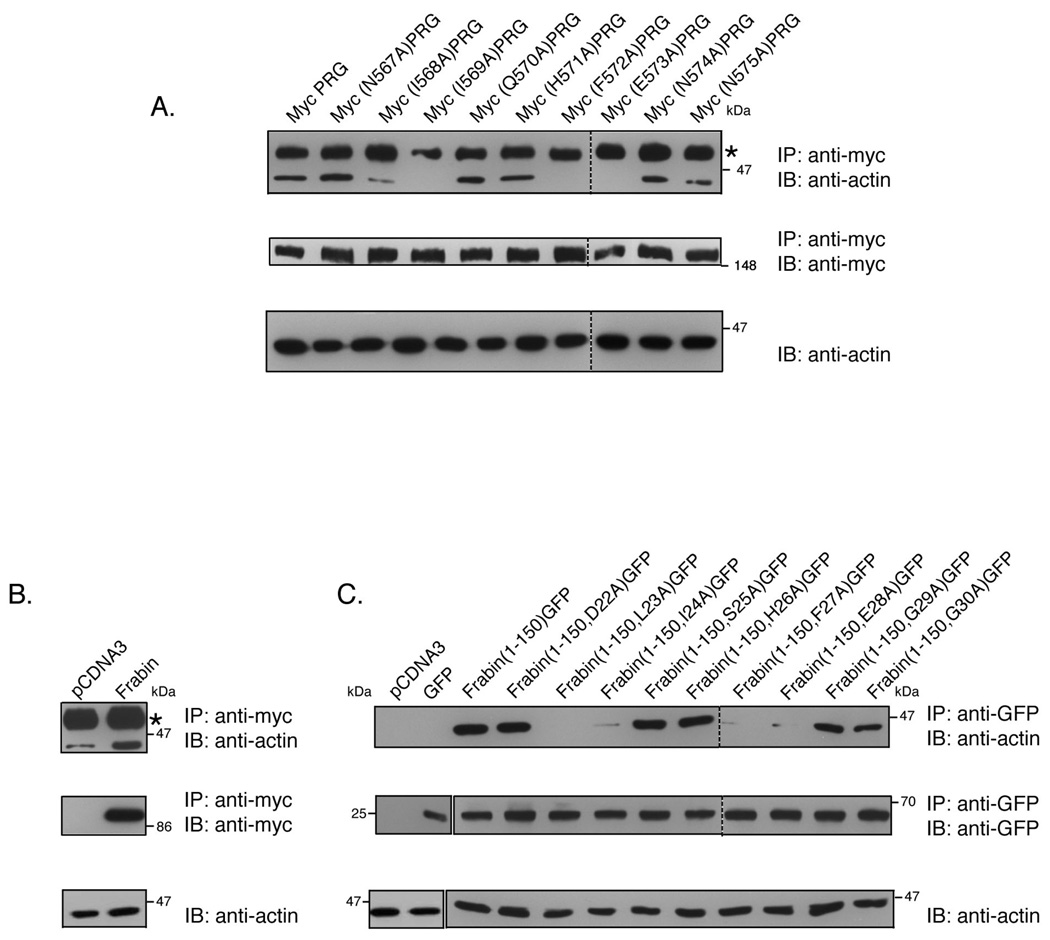
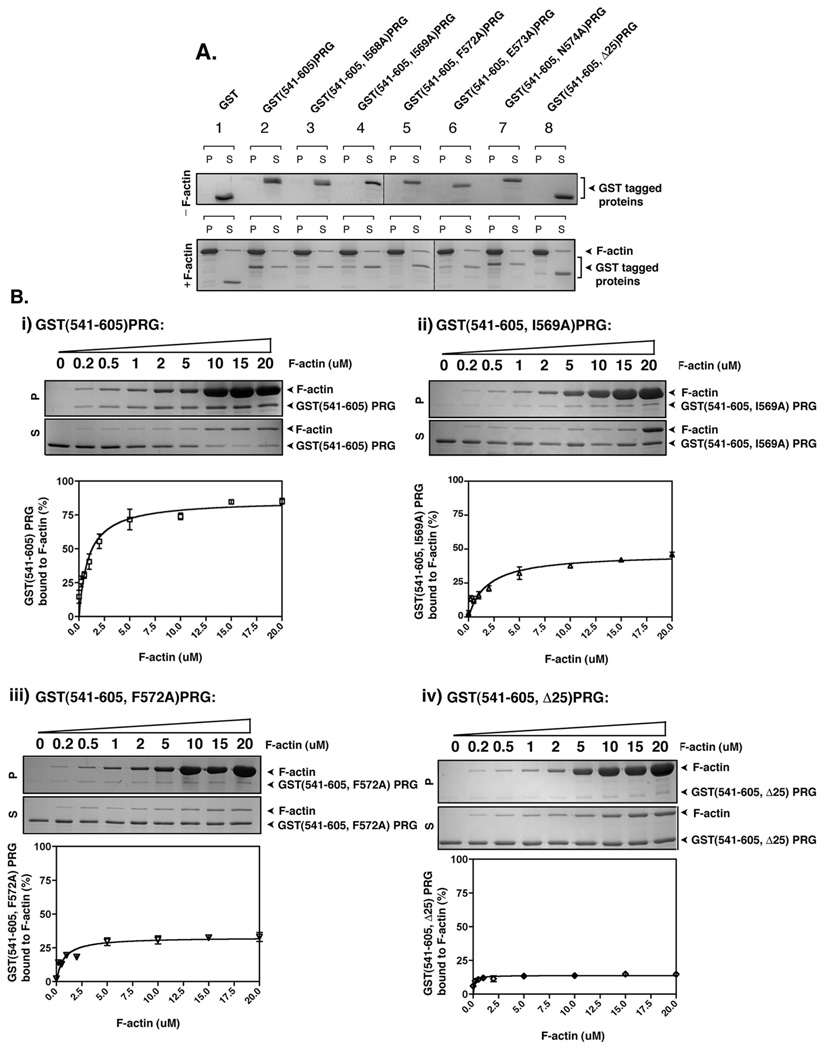
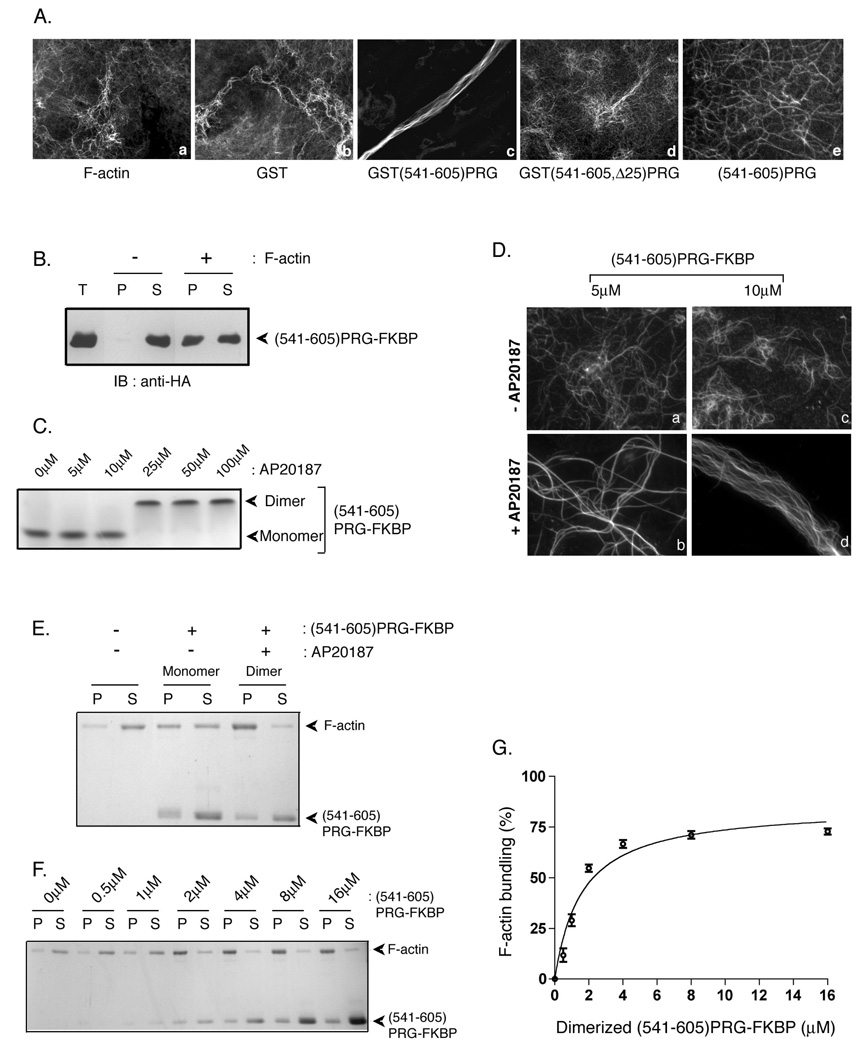
PDZ-RhoGEF is a member of the regulator family of G protein signaling (RGS) domain-containing RhoGEFs (RGS-RhoGEFs) that link activated heterotrimeric G protein alpha subunits of the G12 family to activation of the small GTPase RhoA. Unique among the RGS-RhoGEFs, PDZ-RhoGEF contains a short sequence that localizes the protein to the actin cytoskeleton. In this report, we demonstrate that the actin-binding domain, located between amino acids 561 and 585, directly binds to F-actin in vitro. Extensive mutagenesis identifies isoleucine 568, isoleucine 569, phenylalanine 572, and glutamic acid 573 as being necessary for binding to actin and for colocalization with the actin cytoskeleton in cells. These results define a novel actin-binding sequence in PDZ-RhoGEF with a critical amino acid motif of IIxxFE. Moreover, sequence analysis identifies a similar actin-binding motif in the N-terminus of the RhoGEF frabin, and as with PDZ-RhoGEF, mutagenesis and actin interaction experiments demonstrate an LIxxFE motif, consisting of the key amino acids leucine 23, isoleucine 24, phenylalanine 27, and glutamic acid 28. Taken together, results with PDZ-RhoGEF and frabin identify a novel actin-binding sequence. Lastly, inducible dimerization of the actin-binding region of PDZ-RhoGEF revealed a dimerization-dependent actin bundling activity in vitro. PDZ-RhoGEF exists in cells as a dimer, raising the possibility that PDZ-RhoGEF could influence actin structure in a manner independent of its ability to activate RhoA.
Figures
Similar articles
-
Identification of a novel sequence in PDZ-RhoGEF that mediates interaction with the actin cytoskeleton.Mol Biol Cell. 2004 Apr;15(4):1760-75. doi: 10.1091/mbc.e03-07-0527. Epub 2004 Jan 23. Mol Biol Cell. 2004. PMID: 14742719 Free PMC article.
-
Leukemia-associated Rho guanine nucleotide exchange factor promotes G alpha q-coupled activation of RhoA.Mol Cell Biol. 2002 Jun;22(12):4053-61. doi: 10.1128/MCB.22.12.4053-4061.2002. Mol Cell Biol. 2002. PMID: 12024019 Free PMC article.
-
Identification of a novel actin-binding domain within the Rho guanine nucleotide exchange factor TEM4.PLoS One. 2012;7(7):e41876. doi: 10.1371/journal.pone.0041876. Epub 2012 Jul 24. PLoS One. 2012. PMID: 22911862 Free PMC article.
-
Frabin and other related Cdc42-specific guanine nucleotide exchange factors couple the actin cytoskeleton with the plasma membrane.J Cell Mol Med. 2008 Aug;12(4):1169-76. doi: 10.1111/j.1582-4934.2008.00345.x. Epub 2008 Apr 9. J Cell Mol Med. 2008. PMID: 18410521 Free PMC article. Review.
-
The GAPs, GEFs, and GDIs of heterotrimeric G-protein alpha subunits.Int J Biol Sci. 2005;1(2):51-66. doi: 10.7150/ijbs.1.51. Epub 2005 Apr 1. Int J Biol Sci. 2005. PMID: 15951850 Free PMC article. Review.
Cited by
-
Identification of F-actin as the dynamic hub in a microbial-induced GTPase polarity circuit.Cell. 2012 Feb 17;148(4):803-15. doi: 10.1016/j.cell.2011.11.063. Cell. 2012. PMID: 22341450 Free PMC article.
-
Solo regulates the localization and activity of PDZ-RhoGEF for actin cytoskeletal remodeling in response to substrate stiffness.Mol Biol Cell. 2024 Jun 1;35(6):ar87. doi: 10.1091/mbc.E23-11-0421. Epub 2024 Apr 24. Mol Biol Cell. 2024. PMID: 38656797 Free PMC article.
-
Regulated localization is sufficient for hormonal control of regulator of G protein signaling homology Rho guanine nucleotide exchange factors (RH-RhoGEFs).J Biol Chem. 2014 Jul 11;289(28):19737-46. doi: 10.1074/jbc.M114.564930. Epub 2014 May 22. J Biol Chem. 2014. PMID: 24855647 Free PMC article.
-
Lowe syndrome-linked endocytic adaptors direct membrane cycling kinetics with OCRL in Dictyostelium discoideum.Mol Biol Cell. 2019 Aug 1;30(17):2268-2282. doi: 10.1091/mbc.E18-08-0510. Epub 2019 Jun 19. Mol Biol Cell. 2019. PMID: 31216233 Free PMC article.
-
Systems analysis of RhoGEF and RhoGAP regulatory proteins reveals spatially organized RAC1 signalling from integrin adhesions.Nat Cell Biol. 2020 Apr;22(4):498-511. doi: 10.1038/s41556-020-0488-x. Epub 2020 Mar 23. Nat Cell Biol. 2020. PMID: 32203420
References
-
- Wennerberg K, Rossman KL, Der CJ. The Ras superfamily at a glance. J Cell Sci. 2005;118:843–846. - PubMed
-
- Ridley AJ, Hall A. The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell. 1992;70:389–399. - PubMed
-
- Ridley AJ, Paterson HF, Johnston CL, Diekmann D, Hall A. The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell. 1992;70:401–410. - PubMed
-
- Nobes CD, Hall A. Rho, rac, and cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers,lamellipodia, and filopodia. Cell. 1995;81:53–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
