

Evidence for the direct interaction of spermine with the inwardly rectifying potassium channel
- PMID: 19620244
- PMCID: PMC2758011
- DOI: 10.1074/jbc.M109.029355
Evidence for the direct interaction of spermine with the inwardly rectifying potassium channel
Abstract
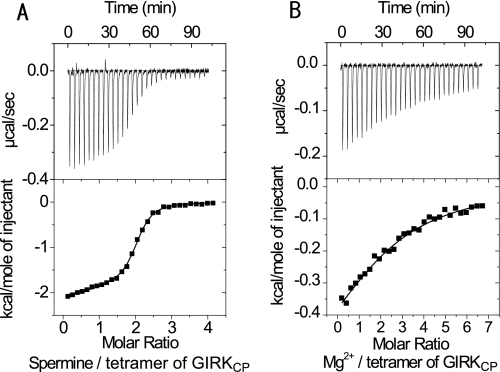
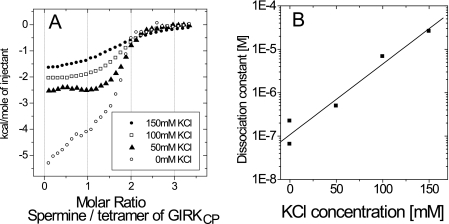
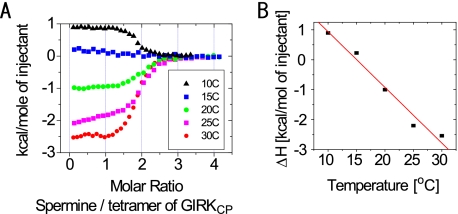
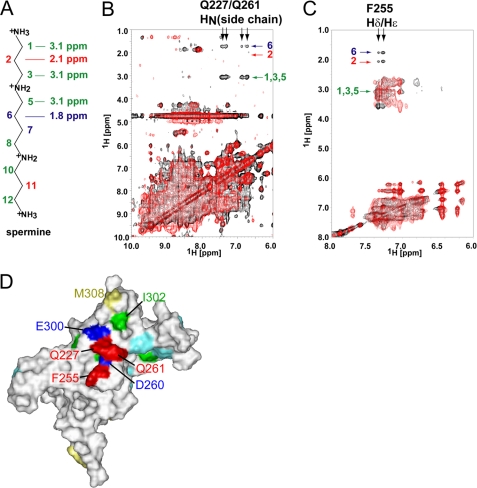
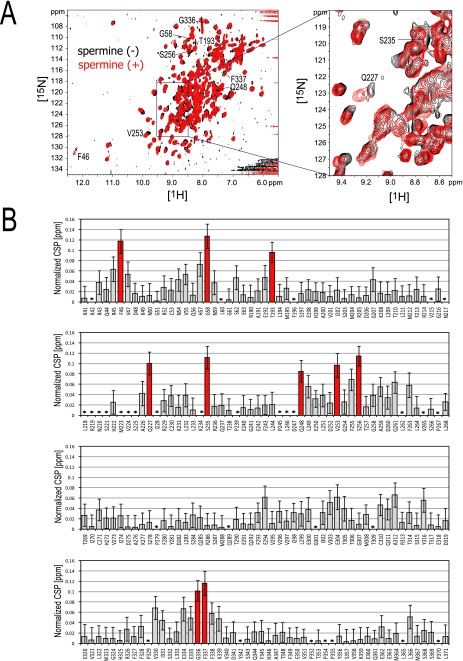
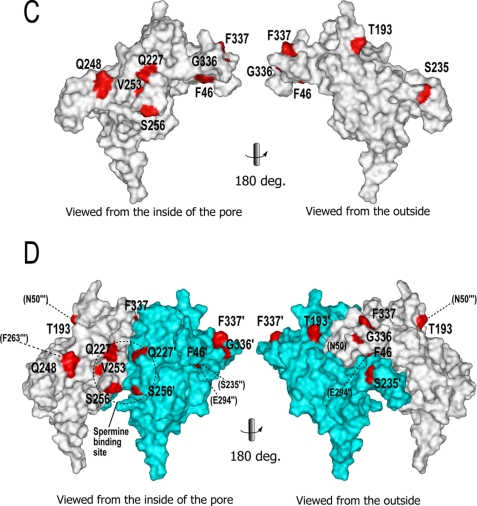
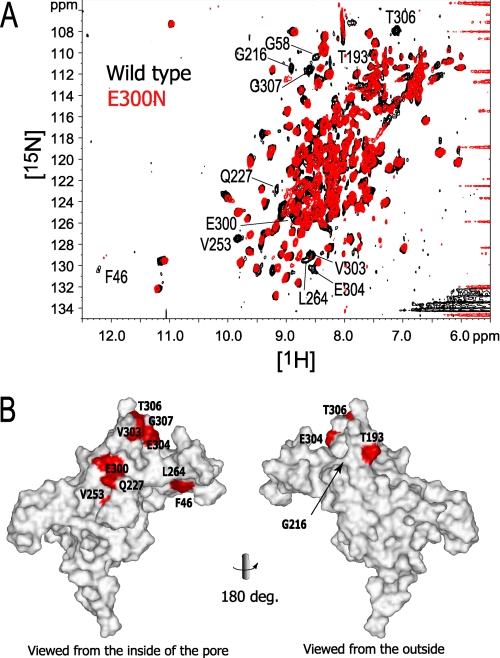
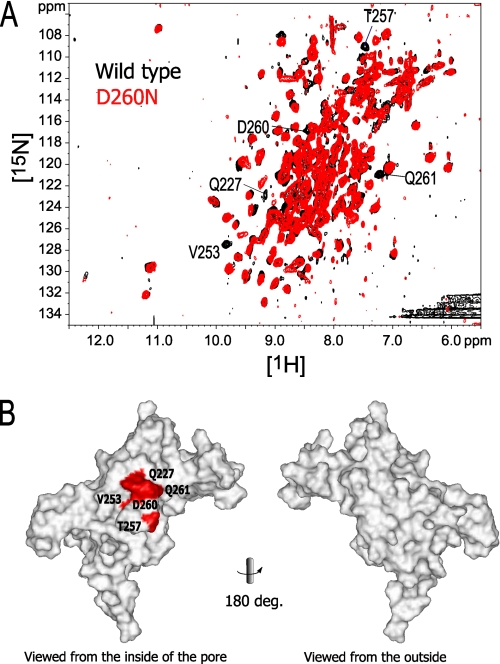
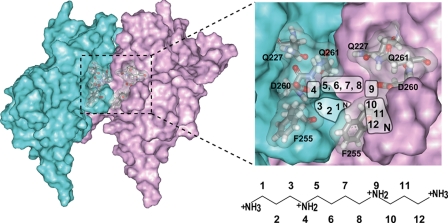
The inwardly rectifying potassium channel (Kir) regulates resting membrane potential, K+ homeostasis, heart rate, and hormone secretion. The outward current is blocked in a voltage-dependent manner, upon the binding of intracellular polyamines or Mg2+ to the transmembrane pore domain. Meanwhile, electrophysiological studies have shown that mutations of several acidic residues in the intracellular regions affected the inward rectification. Although these acidic residues are assumed to bind polyamines, the functional role of the binding of polyamines and Mg2+ to the intracellular regions of Kirs remains unclear. Here, we report thermodynamic and structural studies of the interaction between polyamines and the cytoplasmic pore of mouse Kir3.1/GIRK1, which is gated by binding of G-protein betagamma-subunit (Gbetagamma). ITC analyses showed that two spermine molecules bind to a tetramer of Kir3.1/GIRK1 with a dissociation constant of 26 microM, which is lower than other blockers. NMR analyses revealed that the spermine binding site is Asp-260 and its surrounding area. Small but significant chemical shift perturbations upon spermine binding were observed in the subunit-subunit interface of the tetramer, suggesting that spermine binding alters the relative orientations of the four subunits. Our ITC and NMR results postulated a spermine binding mode, where one spermine molecule bridges two Asp-260 side chains from adjacent subunits, with rearrangement of the subunit orientations. This suggests the functional roles of spermine binding to the cytoplasmic pore: stabilization of the resting state conformation of the channel, and instant translocation to the transmembrane pore upon activation through the Gbetagamma-induced conformational rearrangement.
Figures
References
-
- Bichet D., Haass F. A., Jan L. Y. (2003) Nat. Rev. Neurosci. 4, 957–967 - PubMed
-
- Ficker E., Taglialatela M., Wible B. A., Henley C. M., Brown A. M. (1994) Science 266, 1068–1072 - PubMed
-
- Lopatin A. N., Makhina E. N., Nichols C. G. (1994) Nature 372, 366–369 - PubMed
-
- Fakler B., Brändle U., Glowatzki E., Weidemann S., Zenner H. P., Ruppersberg J. P. (1995) Cell 80, 149–154 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
