

Incorporation of NKT cell-activating glycolipids enhances immunogenicity and vaccine efficacy of Mycobacterium bovis bacillus Calmette-Guerin
- PMID: 19620317
- PMCID: PMC2719834
- DOI: 10.4049/jimmunol.0900858
Incorporation of NKT cell-activating glycolipids enhances immunogenicity and vaccine efficacy of Mycobacterium bovis bacillus Calmette-Guerin
Abstract
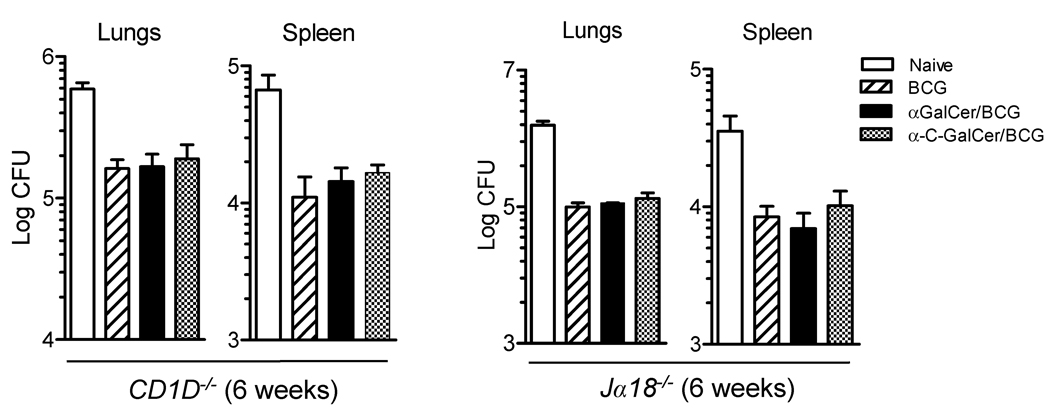
The attenuated strain of Mycobacterium bovis known as bacille Calmette-Guérin (BCG) has been widely used as a vaccine for prevention of disease by Mycobacterium tuberculosis, but with relatively little evidence of success. Recent studies suggest that the failure of BCG may be due to its retention of immune evasion mechanisms that delay or prevent the priming of robust protective cell-mediated immunity. In this study, we describe an approach to enhance the immunogenicity of BCG by incorporating glycolipid activators of CD1d-restricted NKT cells, a conserved T cell subset with the potential to augment many types of immune responses. A method was developed for stably incorporating two forms of the NKT cell activator alpha-galactosylceramide into live BCG organisms, and the impact of this on stimulation of T cell responses and protective antimycobacterial immunity was evaluated. We found that live BCG containing relatively small amounts of incorporated alpha-galactosylceramide retained the ability to robustly activate NKT cells. Compared with immunization with unmodified BCG, the glycolipid-modified BCG stimulated increased maturation of dendritic cells and markedly augmented the priming of Ag-specific CD8(+) T cells responses. These effects were correlated with improved protective effects of vaccination in mice challenged with virulent M. tuberculosis. These results support the view that mycobacteria possess mechanisms to avoid stimulation of CD8(+) T cell responses and that such responses contribute significantly to protective immunity against these pathogens. Our findings raise the possibility of a simple modification of BCG that could yield a more effective vaccine for control of tuberculosis.
Figures






Similar articles
-
Improving Mycobacterium bovis bacillus Calmette-Guèrin as a vaccine delivery vector for viral antigens by incorporation of glycolipid activators of NKT cells.PLoS One. 2014 Sep 25;9(9):e108383. doi: 10.1371/journal.pone.0108383. eCollection 2014. PLoS One. 2014. PMID: 25255287 Free PMC article.
-
Is interferon-gamma the right marker for bacille Calmette-Guérin-induced immune protection? The missing link in our understanding of tuberculosis immunology.Clin Exp Immunol. 2012 Sep;169(3):213-9. doi: 10.1111/j.1365-2249.2012.04614.x. Clin Exp Immunol. 2012. PMID: 22861360 Free PMC article. Review.
-
Dendritic cells induce CD4+ and CD8+ T-cell responses to Mycobacterium bovis and M. avium antigens in Bacille Calmette Guérin vaccinated and nonvaccinated cattle.Scand J Immunol. 2000 Sep;52(3):285-91. doi: 10.1046/j.1365-3083.2000.00780.x. Scand J Immunol. 2000. PMID: 10972905
-
Bacille Calmette-Guérin vaccination enhances human gamma delta T cell responsiveness to mycobacteria suggestive of a memory-like phenotype.J Immunol. 1998 Jul 15;161(2):1045-54. J Immunol. 1998. PMID: 9670986 Clinical Trial.
-
CD1d and natural killer T cells in immunity to Mycobacterium tuberculosis.Adv Exp Med Biol. 2013;783:199-223. doi: 10.1007/978-1-4614-6111-1_11. Adv Exp Med Biol. 2013. PMID: 23468111 Review.
Cited by
-
NK1.1+ cells and IL-22 regulate vaccine-induced protective immunity against challenge with Mycobacterium tuberculosis.J Immunol. 2012 Jul 15;189(2):897-905. doi: 10.4049/jimmunol.1102833. Epub 2012 Jun 18. J Immunol. 2012. PMID: 22711885 Free PMC article.
-
Protective effect of a lipid-based preparation from Mycobacterium smegmatis in a murine model of progressive pulmonary tuberculosis.Biomed Res Int. 2014;2014:273129. doi: 10.1155/2014/273129. Epub 2014 Dec 7. Biomed Res Int. 2014. PMID: 25548767 Free PMC article.
-
Natural Killer T Cells and Mucosal-Associated Invariant T Cells in Lung Infections.Front Immunol. 2018 Aug 2;9:1750. doi: 10.3389/fimmu.2018.01750. eCollection 2018. Front Immunol. 2018. PMID: 30116242 Free PMC article. Review.
-
Lipid and glycolipid antigens of CD1d-restricted natural killer T cells.Semin Immunol. 2010 Apr;22(2):68-78. doi: 10.1016/j.smim.2009.10.003. Epub 2009 Nov 27. Semin Immunol. 2010. PMID: 19945296 Free PMC article. Review.
-
A rapid fluorescence-based assay for classification of iNKT cell activating glycolipids.J Am Chem Soc. 2011 Apr 13;133(14):5198-201. doi: 10.1021/ja200070u. Epub 2011 Mar 22. J Am Chem Soc. 2011. PMID: 21425779 Free PMC article.
References
-
- Dye C, Scheele S, Dolin P, Pathania V, Raviglione MC. Consensus statement. Global burden of tuberculosis: estimated incidence, prevalence, and mortality by country. WHO Global Surveillance and Monitoring Project. JAMA. 1999;282:677–686. - PubMed
-
- Hoft DF. Tuberculosis vaccine development: goals, immunological design, and evaluation. Lancet. 2008;372:164–175. - PubMed
-
- Andersen P. Tuberculosis vaccines - an update. Nature Reviews Microbiology. 2007;5:484–U16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
