

Activation of endothelial intrinsic NF-{kappa}B pathway impairs protein C anticoagulation mechanism and promotes coagulation in endotoxemic mice
- PMID: 19620400
- PMCID: PMC2746475
- DOI: 10.1182/blood-2009-02-205914
Activation of endothelial intrinsic NF-{kappa}B pathway impairs protein C anticoagulation mechanism and promotes coagulation in endotoxemic mice
Abstract
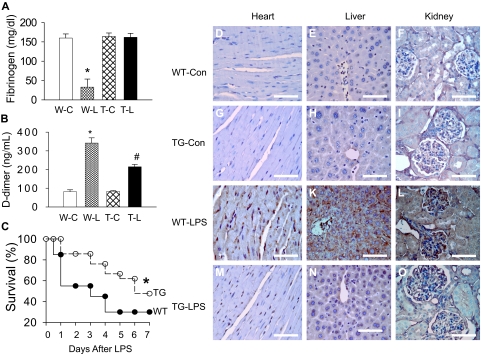
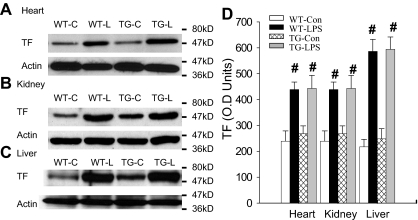
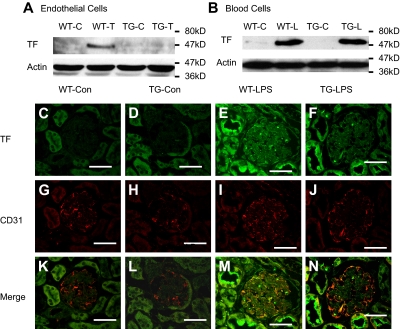
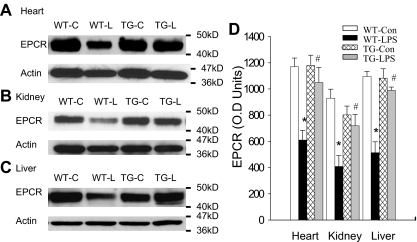
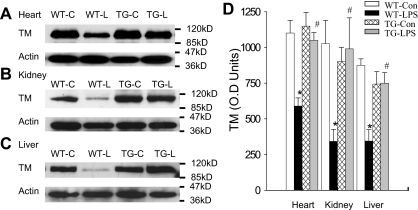
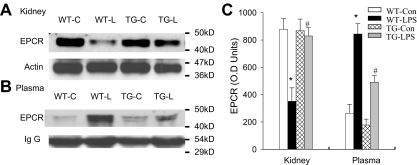
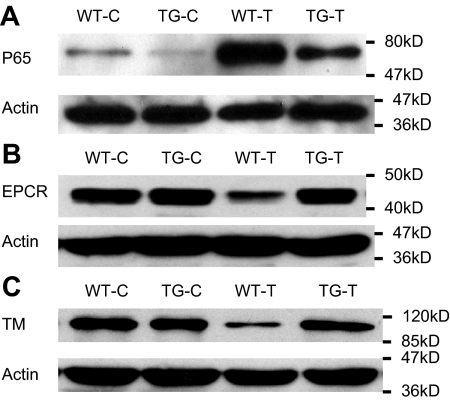
Although the role of systemic activation of the nuclear factor kappaB (NF-kappaB) pathway in septic coagulation has been well documented, little is known about the contribution of endothelial-specific NF-kappaB signaling in this pathologic process. Here, we used transgenic mice that conditionally overexpress a mutant I-kappaBalpha, an inhibitor of NF-kappaB, selectively on endothelium, and their wild-type littermates to define the role of endothelial-specific NF-kappaB in septic coagulation. In wild-type mice, lipopolysaccharide (LPS) challenge (5 mg/kg intraperitoneally) caused markedly increased plasma markers of coagulation, decreased plasma fibrinogen level, and widespread tissue fibrin deposition, which were abrogated by endothelial NF-kappaB blockade in transgenic mice. Endothelial NF-kappaB blockade inhibited tissue factor expression in endothelial cells, but not in leukocytes. Endothelial NF-kappaB blockade did not inhibit LPS-induced tissue factor expression in heart, kidney, and liver. Endothelial NF-kappaB blockade prevented LPS down-regulation of endothelial protein C receptor (EPCR) and thrombomodulin protein expressions, inhibited tissue tumor necrosis factor-alpha converting enzyme activity, reduced EPCR shedding, and restored plasma protein C level. Our data demonstrate that endothelial intrinsic NF-kappaB signaling plays a pivotal role in septic coagulation and suggests a link between endothelial-specific NF-kappaB activation and the impairment of the thrombomodulin-protein C-EPCR anticoagulation pathway.
Figures
References
-
- Martin GS, Mannino DM, Eaton S, Moss M. The epidemiology of sepsis in the United States from 1979 through 2000. N Engl J Med. 2003;348(16):1546–1554. - PubMed
-
- Liu SF, Malik AB. NF-κB activation as a pathologic mechanism of septic shock and inflammation. Am J Physiol Lung Cell Mol Physiol. 2006;290(4):L622–L645. - PubMed
-
- Schouten M, Wiersinga WJ, Levi M, Van Der Poll T. Inflammation, endothelium and coagulation in sepsis. J Leukoc Biol. 2008;83(3):536–545. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
