

Recruitment of functionally distinct membrane proteins to chromatin mediates nuclear envelope formation in vivo
- PMID: 19620630
- PMCID: PMC2717638
- DOI: 10.1083/jcb.200901106
Recruitment of functionally distinct membrane proteins to chromatin mediates nuclear envelope formation in vivo
Erratum in
- J Cell Biol. 2009 Sep 21;186(6):929
Abstract
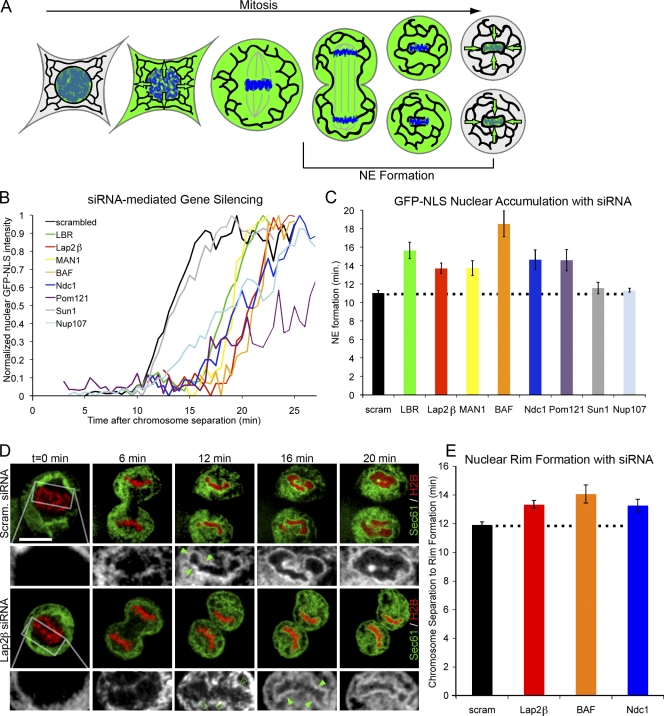
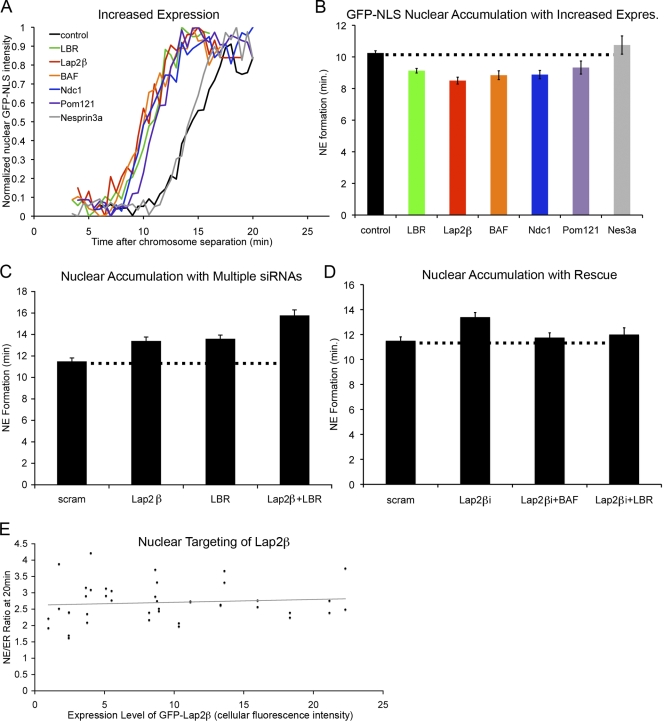
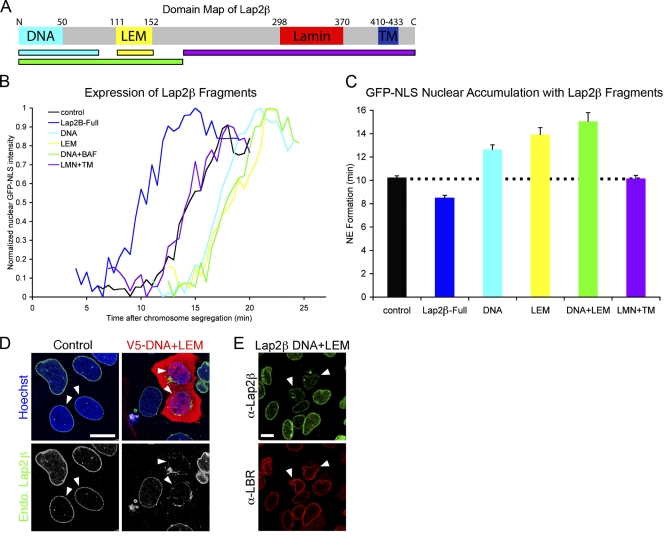
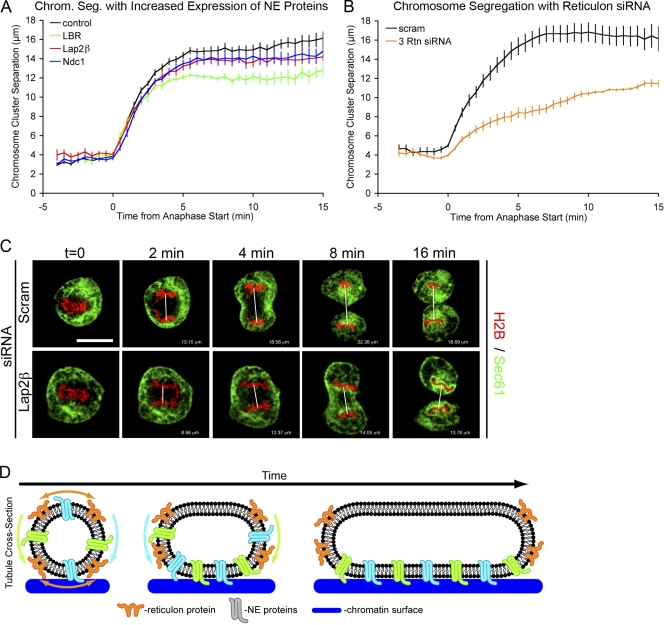
Formation of the nuclear envelope (NE) around segregated chromosomes occurs by the reshaping of the endoplasmic reticulum (ER), a reservoir for disassembled nuclear membrane components during mitosis. In this study, we show that inner nuclear membrane proteins such as lamin B receptor (LBR), MAN1, Lap2beta, and the trans-membrane nucleoporins Ndc1 and POM121 drive the spreading of ER membranes into the emerging NE via their capacity to bind chromatin in a collaborative manner. Despite their redundant functions, decreasing the levels of any of these trans-membrane proteins by RNAi-mediated knockdown delayed NE formation, whereas increasing the levels of any of them had the opposite effect. Furthermore, acceleration of NE formation interferes with chromosome separation during mitosis, indicating that the time frame over which chromatin becomes membrane enclosed is physiologically relevant and regulated. These data suggest that functionally distinct classes of chromatin-interacting membrane proteins, which are present at nonsaturating levels, collaborate to rapidly reestablish the nuclear compartment at the end of mitosis.
Figures
References
-
- Anderson D.J., Hetzer M.W. 2007. Nuclear envelope formation by chromatin-mediated reorganization of the endoplasmic reticulum.Nat. Cell Biol. 9:1160–1166 - PubMed
-
- Anderson D.J., Hetzer M.W. 2008b. Shaping the endoplasmic reticulum into the nuclear envelope.J. Cell Sci. 121:137–142 - PubMed
-
- Antonin W., Ellenberg J., Dultz E. 2008. Nuclear pore complex assembly through the cell cycle: regulation and membrane organization.FEBS Lett. 582:2004–2016 - PubMed
-
- Burke B., Ellenberg J. 2002. Remodelling the walls of the nucleus.Nat. Rev. Mol. Cell Biol. 3:487–497 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
