

Prolylcarboxypeptidase regulates food intake by inactivating alpha-MSH in rodents
- PMID: 19620781
- PMCID: PMC2719925
- DOI: 10.1172/JCI37209
Prolylcarboxypeptidase regulates food intake by inactivating alpha-MSH in rodents
Abstract
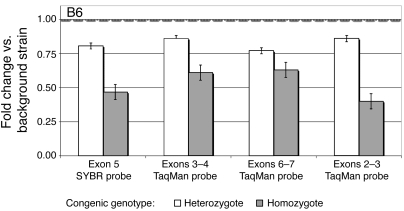
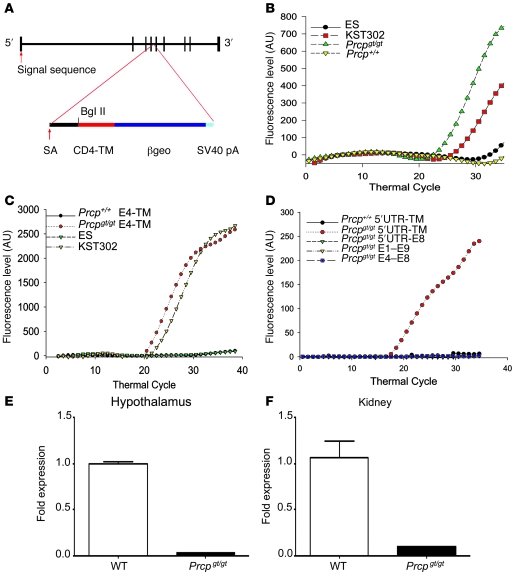
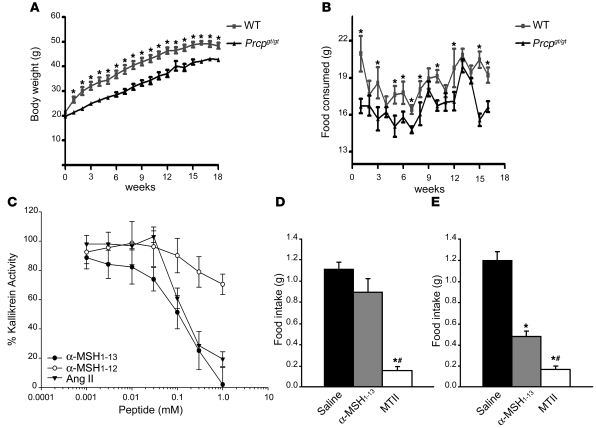
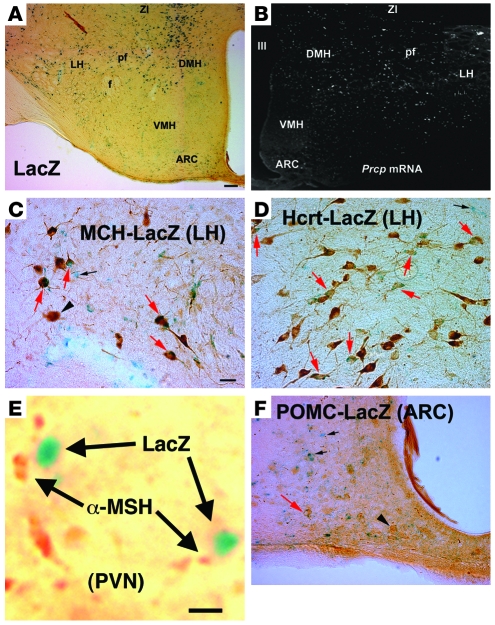
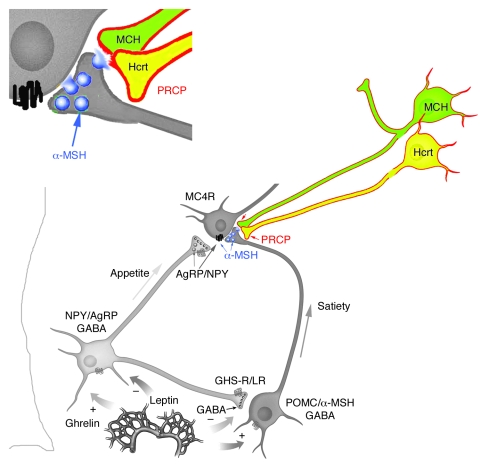
The anorexigenic neuromodulator alpha-melanocyte-stimulating hormone (alpha-MSH; referred to here as alpha-MSH1-13) undergoes extensive posttranslational processing, and its in vivo activity is short lived due to rapid inactivation. The enzymatic control of alpha-MSH1-13 maturation and inactivation is incompletely understood. Here we have provided insight into alpha-MSH1-13 inactivation through the generation and analysis of a subcongenic mouse strain with reduced body fat compared with controls. Using positional cloning, we identified a maximum of 6 coding genes, including that encoding prolylcarboxypeptidase (PRCP), in the donor region. Real-time PCR revealed a marked genotype effect on Prcp mRNA expression in brain tissue. Biochemical studies using recombinant PRCP demonstrated that PRCP removes the C-terminal amino acid of alpha-MSH1-13, producing alpha-MSH1-12, which is not neuroactive. We found that Prcp was expressed in the hypothalamus in neuronal populations that send efferents to areas where alpha-MSH1-13 is released from axon terminals. The inhibition of PRCP activity by small molecule protease inhibitors administered peripherally or centrally decreased food intake in both wild-type and obese mice. Furthermore, Prcp-null mice had elevated levels of alpha-MSH1-13 in the hypothalamus and were leaner and shorter than the wild-type controls on a regular chow diet; they were also resistant to high-fat diet-induced obesity. Our results suggest that PRCP is an important component of melanocortin signaling and weight maintenance via control of active alpha-MSH1-13 levels.
Figures


Comment in
-
Reduced levels of neurotransmitter-degrading enzyme PRCP promote a lean phenotype. [corrected].J Clin Invest. 2009 Aug;119(8):2130-3. doi: 10.1172/JCI40001. Epub 2009 Jul 20. J Clin Invest. 2009. PMID: 19620779 Free PMC article.
References
-
- Stylianou I.M., et al. Microarray gene expression analysis of the Fob3b obesity QTL identifies positional candidate gene Sqle and perturbed cholesterol and glycolysis pathways. Physiol. Genomics. 2005;20:224–232. - PubMed
-
- Williams D.L., Schwartz M.W. The melanocortin system as a central integrator of direct and indirect controls of food intake. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005;289:R2–R3. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK080003/DK/NIDDK NIH HHS/United States
- DK70039/DK/NIDDK NIH HHS/United States
- DK52581/DK/NIDDK NIH HHS/United States
- R01 HL052779/HL/NHLBI NIH HHS/United States
- HL65194/HL/NHLBI NIH HHS/United States
- P01 HL057346/HL/NHLBI NIH HHS/United States
- DK080003/DK/NIDDK NIH HHS/United States
- P20RR021929/RR/NCRR NIH HHS/United States
- R01 DK084065/DK/NIDDK NIH HHS/United States
- P20 RR021929/RR/NCRR NIH HHS/United States
- R01 DK070039/DK/NIDDK NIH HHS/United States
- R01 HL065194/HL/NHLBI NIH HHS/United States
- R01 DK052581/DK/NIDDK NIH HHS/United States
- HL52779/HL/NHLBI NIH HHS/United States
- HL57346/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
