

The muscle-specific microRNA miR-206 blocks human rhabdomyosarcoma growth in xenotransplanted mice by promoting myogenic differentiation
- PMID: 19620785
- PMCID: PMC2719932
- DOI: 10.1172/JCI38075
The muscle-specific microRNA miR-206 blocks human rhabdomyosarcoma growth in xenotransplanted mice by promoting myogenic differentiation
Abstract
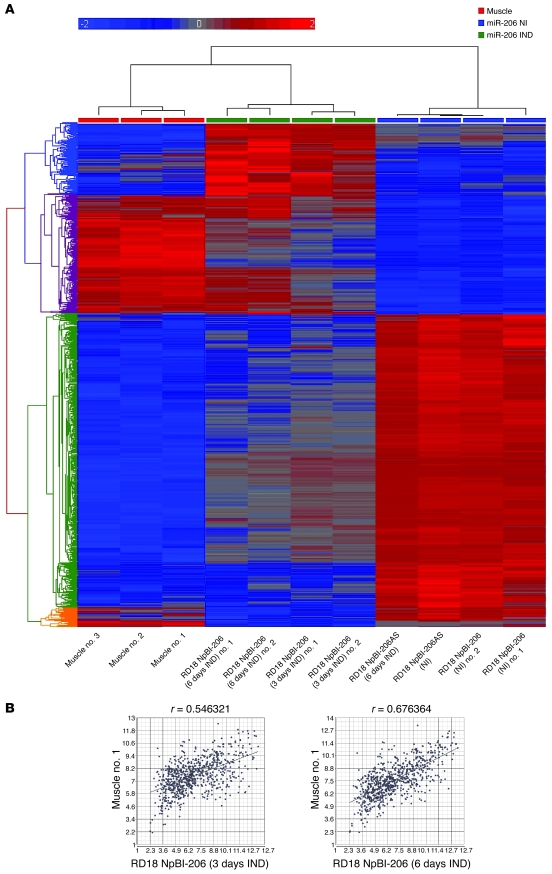
Many microRNAs (miRNAs), posttranscriptional regulators of numerous cellular processes and developmental events, are downregulated in tumors. However, their role in tumorigenesis remains largely unknown. In this work, we examined the role of the muscle-specific miRNAs miR-1 and miR-206 in human rhabdomyosarcoma (RMS), a soft tissue sarcoma thought to arise from skeletal muscle progenitors. We have shown that miR-1 was barely detectable in primary RMS of both the embryonal and alveolar subtypes and that both miR-1 and miR-206 failed to be induced in RMS cell lines upon serum deprivation. Moreover, reexpression of miR-206 in RMS cells promoted myogenic differentiation and blocked tumor growth in xenografted mice by switching the global mRNA expression profile to one that resembled mature muscle. Finally, we showed that the product of the MET proto-oncogene, the Met tyrosine-kinase receptor, which is overexpressed in RMS and has been implicated in RMS pathogenesis, was downregulated in murine satellite cells by miR-206 at the onset of normal myogenesis. Thus, failure of posttranscriptional modulation may underlie Met overexpression in RMS and other types of cancer. We propose that tissue-specific miRNAs such as miR-1 and miR-206, given their ability to modulate hundreds of transcripts and to act as nontoxic differentiating agents, may override the genomic heterogeneity of solid tumors and ultimately hold greater therapeutic potential than single gene-directed drugs.
Figures





Comment in
-
MicroRNA reexpression as differentiation therapy in cancer.J Clin Invest. 2009 Aug;119(8):2119-23. doi: 10.1172/JCI40107. Epub 2009 Jul 20. J Clin Invest. 2009. PMID: 19620782 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
