

Prostaglandin E2, Wnt, and BMP in gastric tumor mouse models
- PMID: 19622104
- PMCID: PMC11159255
- DOI: 10.1111/j.1349-7006.2009.01258.x
Prostaglandin E2, Wnt, and BMP in gastric tumor mouse models
Abstract
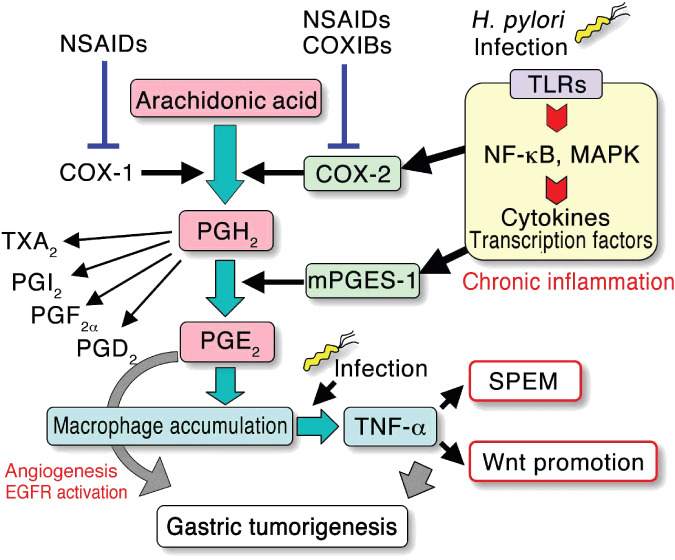
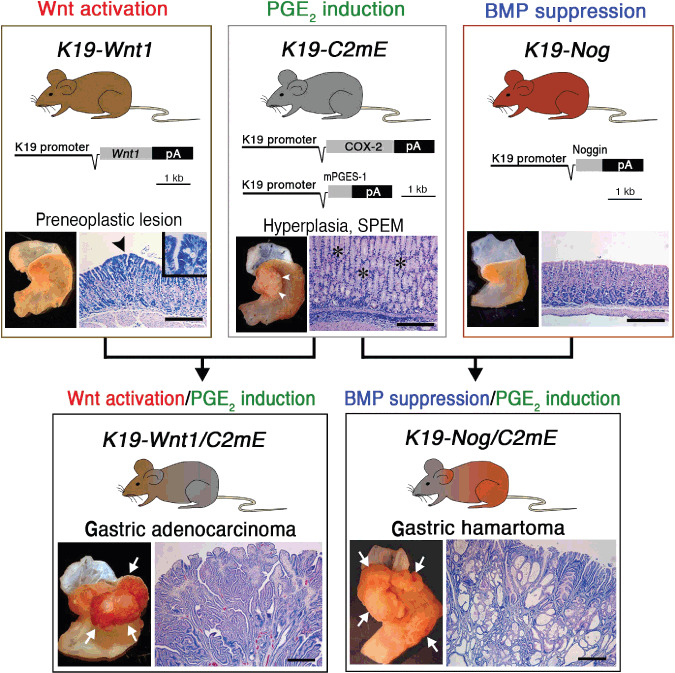
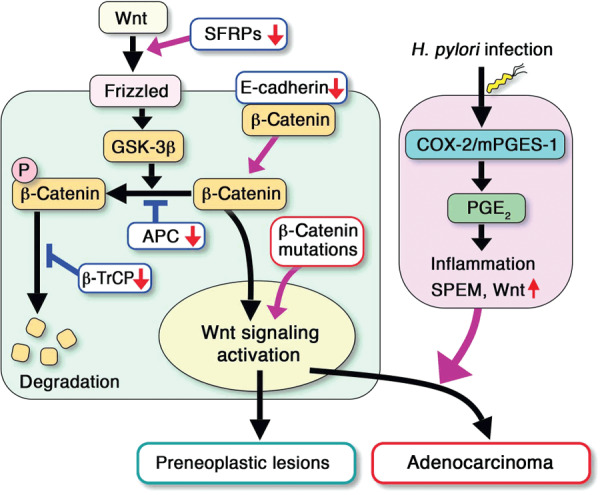
The development of gastric cancer is closely associated with Helicobacter pylori (H. pylori) infection. The expression of cylooxigenase-2 (COX-2), a rate-limiting enzyme for prostaglandin biosynthesis, is induced in H. pylori-associated chronic gastritis, which thus results in the induction of proinflammatory prostaglandin, PGE(2). The COX-2/PGE(2) pathway plays a key role in gastric tumorigenesis. On the other hand, several oncogenic pathways have been shown to trigger gastric tumorigenesis. The activation of Wnt/beta-catenin signaling is found in 30-50% of gastric cancers, thus suggesting that Wnt signaling plays a causal role in gastric cancer development. Mutations in the bone morphogenetic protein (BMP) signaling pathway are responsible for the subset of juvenile polyposis syndrome (JPS) that develops hamartomas in the gastrointestinal tract. BMP suppression appears to contribute to gastric cancer development because gastric cancer risk is increased in JPS. Wnt signaling is important for the maintenance of gastrointestinal stem cells, while BMP promotes epithelial cell differentiation. Accordingly, it is possible that both Wnt activation and BMP suppression can cause gastric tumorigenesis through enhancement of the undifferentiated status of epithelial cells. Recent mouse model studies have indicated that induction of the PGE(2) pathway is required for the development of both gastric adenocarcinoma and hamartoma in the Wnt-activated and BMP-suppressed gastric mucosa, respectively. This article reviews the involvement of the PGE(2), Wnt, and BMP pathways in the development of gastric cancer, and gastric phenotypes that are found in transgenic mouse models of PGE(2) induction, Wnt activation, BMP suppression, or a combination of these pathways.
Figures

References
-
- Thun MJ, Namboodiri MM, Heath CW Jr. Aspirin use and reduced risk of fatal colon cancer. N Engl J Med 1991; 325: 1593–6. - PubMed
-
- Fletcher BS, Kujubu DA, Perrin DM, Herschman HR. Structure of the mitogen‐inducible TIS10 gene and demonstration that the TIS10‐encoded protein is a functional prostaglandin G/H synthase. J Biol Chem 1992; 267: 4338–44. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
