

Role for RACK1 orthologue Cpc2 in the modulation of stress response in fission yeast
- PMID: 19625445
- PMCID: PMC2743619
- DOI: 10.1091/mbc.e09-05-0388
Role for RACK1 orthologue Cpc2 in the modulation of stress response in fission yeast
Abstract
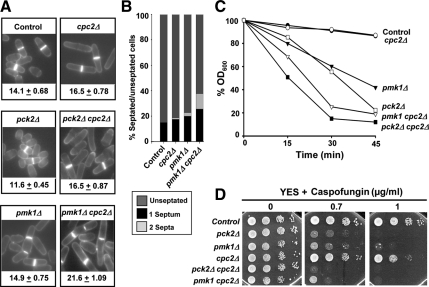
The receptor of activated C kinase (RACK1) is a protein highly conserved among eukaryotes. In mammalian cells, RACK1 functions as an adaptor to favor protein kinase C (PKC)-mediated phosphorylation and subsequent activation of c-Jun NH(2)-terminal kinase mitogen-activated protein kinase. Cpc2, the RACK1 orthologue in the fission yeast Schizosaccharomyces pombe, is involved in the control of G2/M transition and interacts with Pck2, a PKC-type protein member of the cell integrity Pmk1 mitogen-activated protein kinase (MAPK) pathway. Both RACK1 and Cpc2 are structural components of the 40S ribosomal subunit, and recent data suggest that they might be involved in the control of translation. In this work, we present data supporting that Cpc2 negatively regulates the cell integrity transduction pathway by favoring translation of the tyrosine-phosphatases Pyp1 and Pyp2 that deactivate Pmk1. In addition, Cpc2 positively regulates the synthesis of the stress-responsive transcription factor Atf1 and the cytoplasmic catalase, a detoxificant enzyme induced by treatment with hydrogen peroxide. These results provide for the first time strong evidence that the RACK1-type Cpc2 protein controls from the ribosome the extent of the activation of MAPK cascades, the cellular defense against oxidative stress, and the progression of the cell cycle by regulating positively the translation of specific gene products involved in key biological processes.
Figures









Similar articles
-
Fission yeast receptor of activated C kinase (RACK1) ortholog Cpc2 regulates mitotic commitment through Wee1 kinase.J Biol Chem. 2010 Dec 31;285(53):41366-73. doi: 10.1074/jbc.M110.173815. Epub 2010 Oct 25. J Biol Chem. 2010. PMID: 20974849 Free PMC article.
-
Cmk2 kinase is essential for survival in arsenite by modulating translation together with RACK1 orthologue Cpc2 in Schizosaccharomyces pombe.Free Radic Biol Med. 2018 Dec;129:116-126. doi: 10.1016/j.freeradbiomed.2018.09.024. Epub 2018 Sep 18. Free Radic Biol Med. 2018. PMID: 30236788
-
Receptor for activated C-kinase (RACK1) homolog Cpc2 facilitates the general amino acid control response through Gcn2 kinase in fission yeast.J Biol Chem. 2013 Jun 28;288(26):19260-8. doi: 10.1074/jbc.M112.445270. Epub 2013 May 13. J Biol Chem. 2013. PMID: 23671279 Free PMC article.
-
[Cell surface protein Ecm33 is involved in negative feedback regulation of MAP kinase signalling and development of the in vivo real-time monitoring of MAP kinase signalling].Yakugaku Zasshi. 2011;131(8):1195-200. doi: 10.1248/yakushi.131.1195. Yakugaku Zasshi. 2011. PMID: 21804323 Review. Japanese.
-
Phospho-mimicking Atf1 mutants bypass the transcription activating function of the MAP kinase Sty1 of fission yeast.Curr Genet. 2018 Feb;64(1):97-102. doi: 10.1007/s00294-017-0730-7. Epub 2017 Aug 10. Curr Genet. 2018. PMID: 28799013 Review.
Cited by
-
The Receptor for Activated C Kinase in Plant Signaling: Tale of a Promiscuous Little Molecule.Front Plant Sci. 2015 Dec 8;6:1090. doi: 10.3389/fpls.2015.01090. eCollection 2015. Front Plant Sci. 2015. PMID: 26697044 Free PMC article. Review.
-
Fission yeast receptor of activated C kinase (RACK1) ortholog Cpc2 regulates mitotic commitment through Wee1 kinase.J Biol Chem. 2010 Dec 31;285(53):41366-73. doi: 10.1074/jbc.M110.173815. Epub 2010 Oct 25. J Biol Chem. 2010. PMID: 20974849 Free PMC article.
-
RACK1, A multifaceted scaffolding protein: Structure and function.Cell Commun Signal. 2011 Oct 6;9:22. doi: 10.1186/1478-811X-9-22. Cell Commun Signal. 2011. PMID: 21978545 Free PMC article.
-
Genetic relationships between the RACK1 homolog cpc-2 and heterotrimeric G protein subunit genes in Neurospora crassa.PLoS One. 2019 Oct 3;14(10):e0223334. doi: 10.1371/journal.pone.0223334. eCollection 2019. PLoS One. 2019. PMID: 31581262 Free PMC article.
-
A network approach to predict pathogenic genes for Fusarium graminearum.PLoS One. 2010 Oct 4;5(10):e13021. doi: 10.1371/journal.pone.0013021. PLoS One. 2010. PMID: 20957229 Free PMC article.
References
-
- Alfa C., Fantes P., Hyams J., Mcleod M., Warbrick E. Experiments with Fission Yeast. A Laboratory Course Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1993.
-
- Bahler J., Wu J. Q., Longtine M. S., Shah N. G., McKenzie A., 3rd, Steever A. B., Wach A., Philippsen P., Pringle J. R. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 1998;14:943–951. - PubMed
-
- Barba G., Soto T., Madrid M., Núñez A., Vicente J., Gacto M., Cansado J. Activation of the cell integrity pathway is channelled through diverse signalling elements in fission yeast. Cell. Signal. 2008;20:748–757. - PubMed
-
- Benko Z., Sipiczki M., Carr A. M. Cloning of caf1+, caf2+ and caf4+ from Schizosaccharomyces pombe: their involvement in multidrug resistance, UV and pH sensitivity. Mol. Gen. Genet. 1998;260:434–443. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
