

Preerythrocytic, live-attenuated Plasmodium falciparum vaccine candidates by design
- PMID: 19625622
- PMCID: PMC2714279
- DOI: 10.1073/pnas.0906387106
Preerythrocytic, live-attenuated Plasmodium falciparum vaccine candidates by design
Abstract
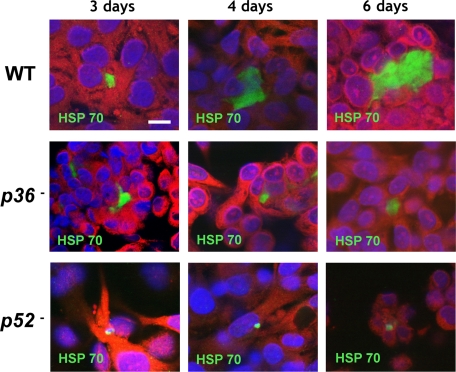
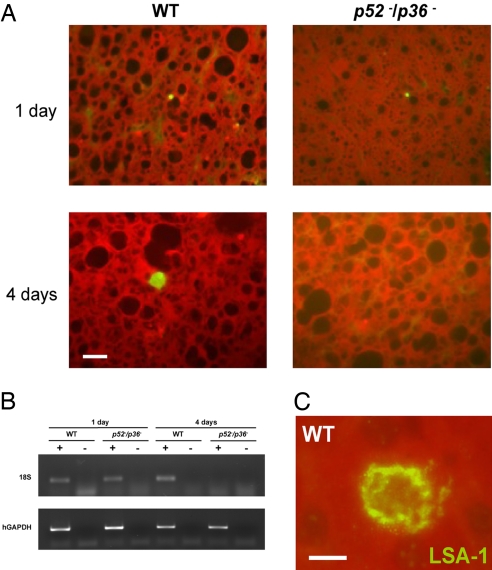
Falciparum malaria is initiated when Anopheles mosquitoes transmit the Plasmodium sporozoite stage during a blood meal. Irradiated sporozoites confer sterile protection against subsequent malaria infection in animal models and humans. This level of protection is unmatched by current recombinant malaria vaccines. However, the live-attenuated vaccine approach faces formidable obstacles, including development of accurate, reproducible attenuation techniques. We tested whether Plasmodium falciparum could be attenuated at the early liver stage by genetic engineering. The P. falciparum genetically attenuated parasites (GAPs) harbor individual deletions or simultaneous deletions of the sporozoite-expressed genes P52 and P36. Gene deletions were done by double-cross-over recombination to avoid genetic reversion of the knockout parasites. The gene deletions did not affect parasite replication throughout the erythrocytic cycle, gametocyte production, mosquito infections, and sporozoite production rates. However, the deletions caused parasite developmental arrest during hepatocyte infection. The double-gene deletion line exhibited a more severe intrahepatocytic growth defect compared with the single-gene deletion lines, and it did not persist. This defect was assessed in an in vitro liver-stage growth assay and in a chimeric mouse model harboring human hepatocytes. The strong phenotype of the double knockout GAP justifies its human testing as a whole-organism vaccine candidate using the established sporozoite challenge model. GAPs might provide a safe and reproducible platform to develop an efficacious whole-cell malaria vaccine that prevents infection at the preerythrocytic stage.
Conflict of interest statement
Conflict of interest statement: S.H.I.K. is an inventor listed on U.S. Patent No. 7,22,179, U.S. Patent No. 7,261,884, and international patent application PCT/US2004/043023, each titled “Live Genetically Attenuated Malaria Vaccine.”
Figures

References
-
- Nussenzweig RS, Vanderberg J, Most H, Orton C. Protective immunity produced by the injection of x-irradiated sporozoites of plasmodium berghei. Nature. 1967;216:160–162. - PubMed
-
- Clyde DF, Most H, McCarthy VC, Vanderberg JP. Immunization of man against sporozite-induced falciparum malaria. Am J Med Sci. 1973;266:169–177. - PubMed
-
- Rieckmann KH, Carson PE, Beaudoin RL, Cassells JS, Sell KW. Letter: Sporozoite induced immunity in man against an Ethiopian strain of Plasmodium falciparum. Trans R Soc Trop Med Hyg. 1974;68:258–259. - PubMed
-
- Hoffman SL, et al. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J Infect Dis. 2002;185:1155–1164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
