

A molecular insight into complement evasion by the staphylococcal complement inhibitor protein family
- PMID: 19625656
- PMCID: PMC2881335
- DOI: 10.4049/jimmunol.0901443
A molecular insight into complement evasion by the staphylococcal complement inhibitor protein family
Abstract
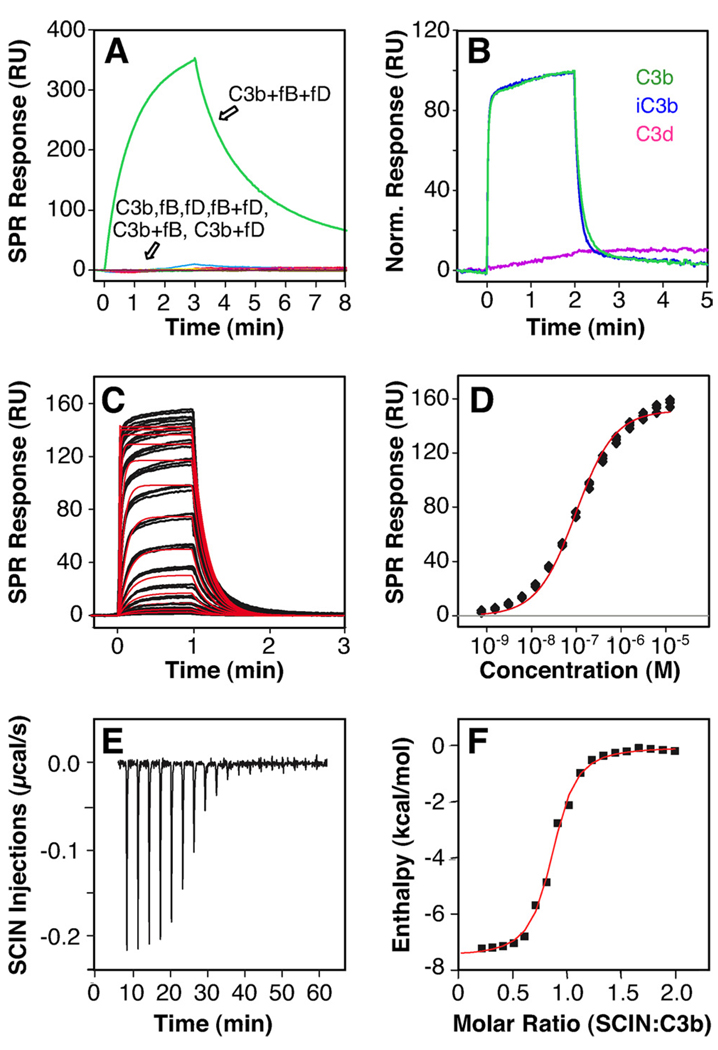
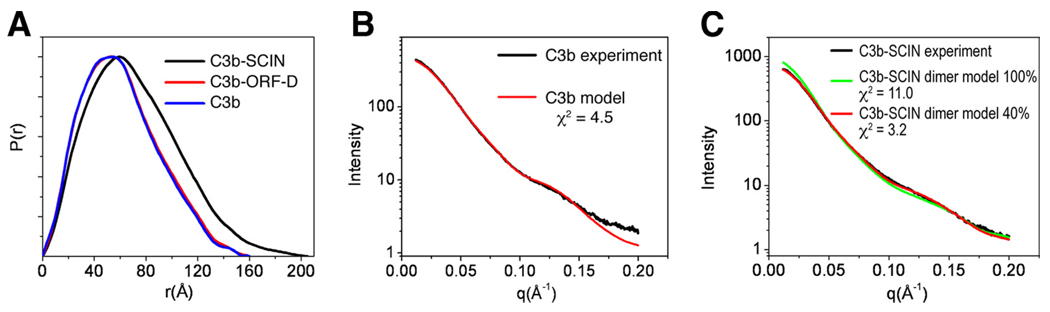
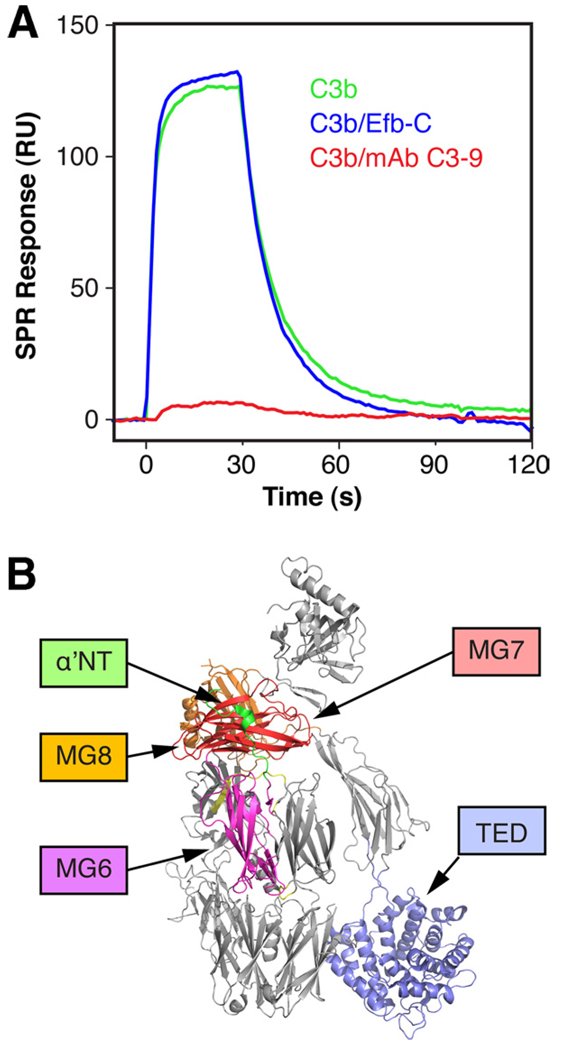
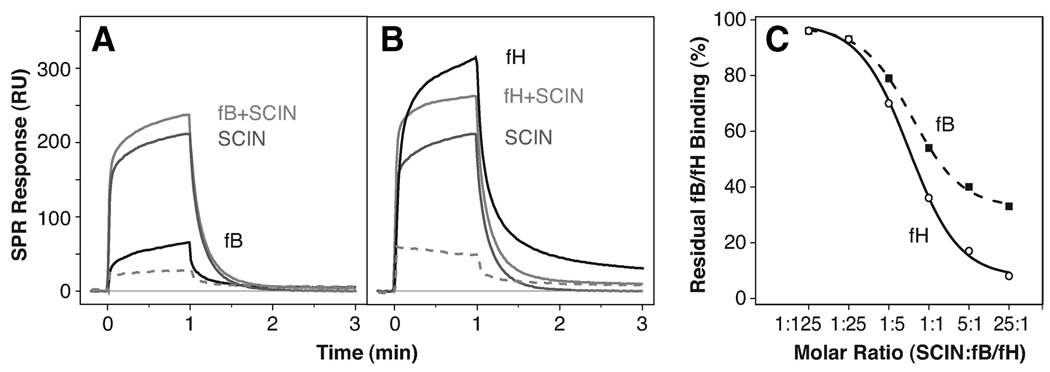
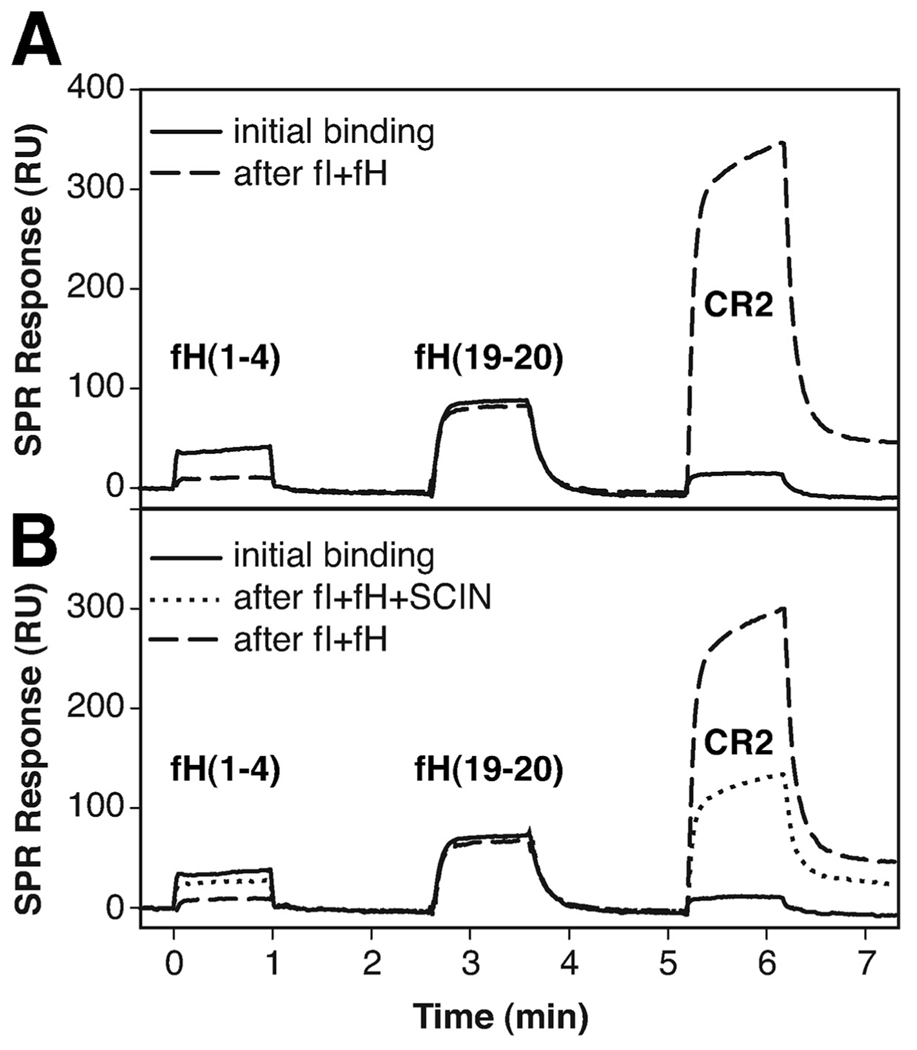
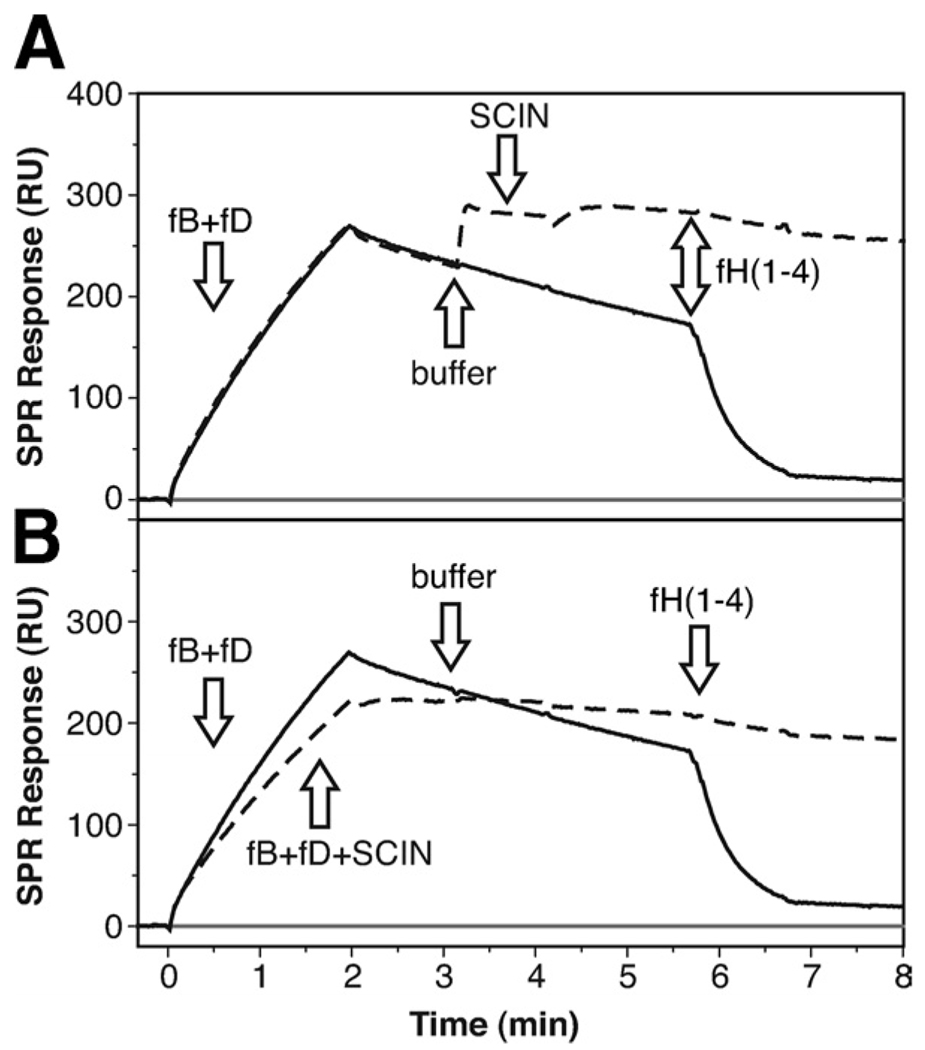
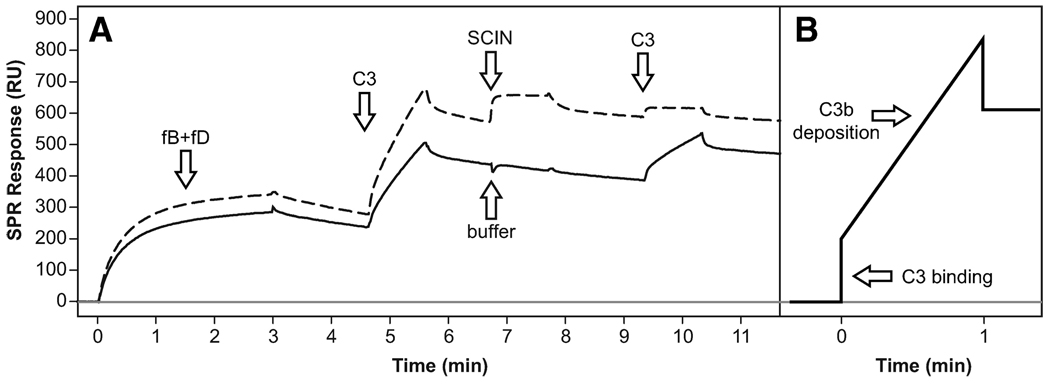
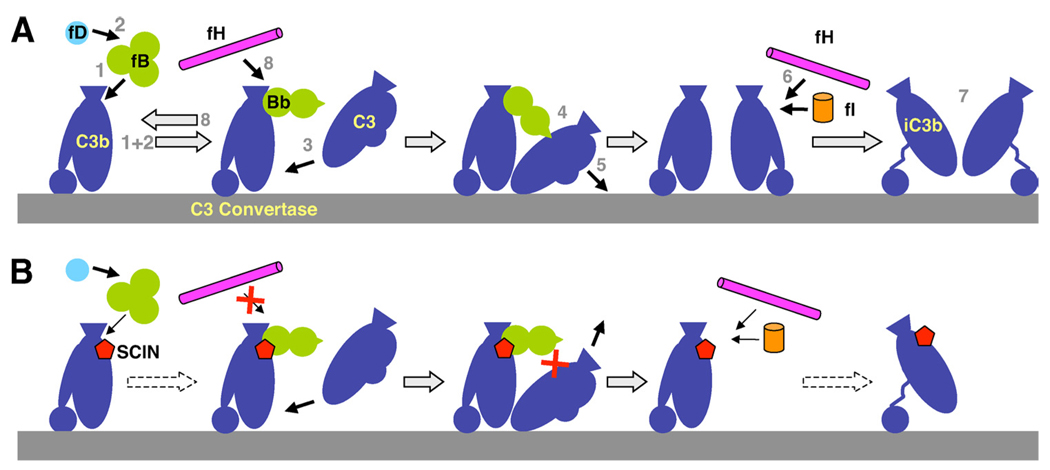
Staphylococcus aureus possesses an impressive arsenal of complement evasion proteins that help the bacterium escape attack of the immune system. The staphylococcal complement inhibitor (SCIN) protein exhibits a particularly high potency and was previously shown to block complement by acting at the level of the C3 convertases. However, many details about the exact binding and inhibitory mechanism remained unclear. In this study, we demonstrate that SCIN directly binds with nanomolar affinity to a functionally important area of C3b that lies near the C terminus of its beta-chain. Direct competition of SCIN with factor B for C3b slightly decreased the formation of surface-bound convertase. However, the main inhibitory effect can be attributed to an entrapment of the assembled convertase in an inactive state. Whereas native C3 is still able to bind to the blocked convertase, no generation and deposition of C3b could be detected in the presence of SCIN. Furthermore, SCIN strongly competes with the binding of factor H to C3b and influences its regulatory activities: the SCIN-stabilized convertase was essentially insensitive to decay acceleration by factor H and the factor I- and H-mediated conversion of surface-bound C3b to iC3b was significantly reduced. By targeting a key area on C3b, SCIN is able to block several essential functions within the alternative pathway, which explains the high potency of the inhibitor. Our findings provide an important insight into complement evasion strategies by S. aureus and may act as a base for further functional studies.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures
References
-
- Lowy FD. Staphylococcus aureus infections. N. Engl. J. Med. 1998;339:520–532. - PubMed
-
- Lee LY, Hook M, Haviland D, Wetsel RA, Yonter EO, Syribeys P, Vernachio J, Brown EL. Inhibition of complement activation by a secreted Staphylococcus aureus protein. J. Infect. Dis. 2004;190:571–579. - PubMed
-
- Chavakis T, Preissner KT, Herrmann M. The anti-inflammatory activities of Staphylococcus aureus. Trends Immunol. 2007;28:408–418. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
