

Limiting transport steps and novel interactions of Connexin-43 along the secretory pathway
- PMID: 19626334
- PMCID: PMC2756399
- DOI: 10.1007/s00418-009-0617-x
Limiting transport steps and novel interactions of Connexin-43 along the secretory pathway
Abstract
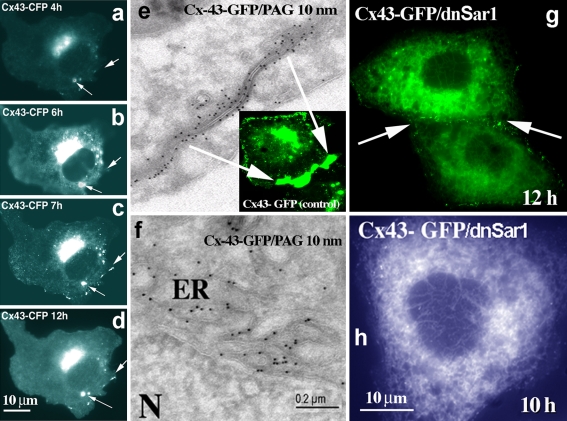
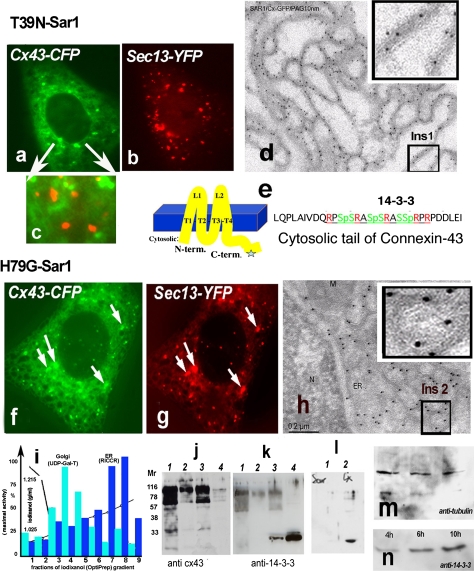
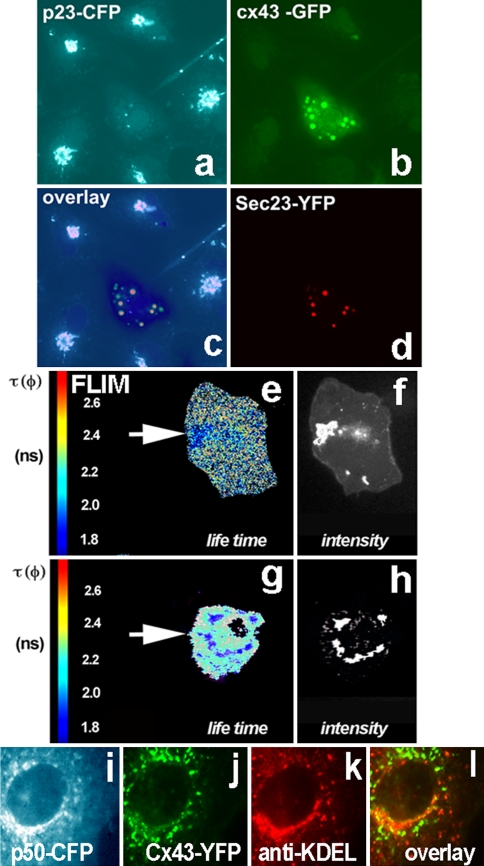
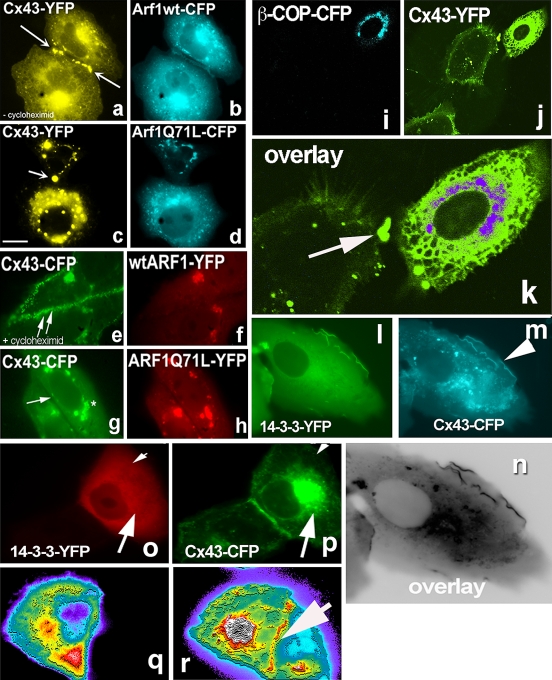
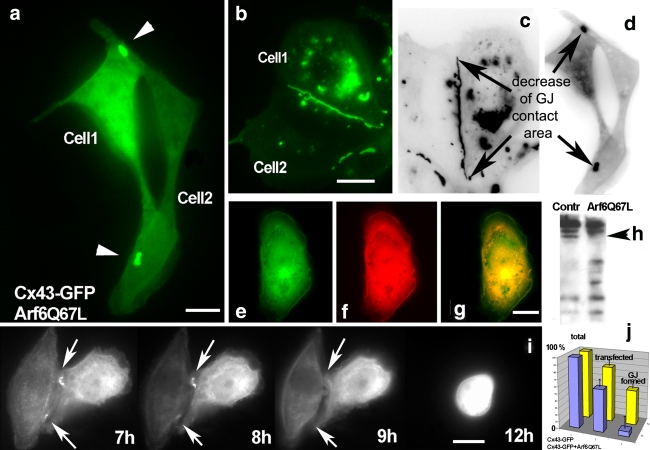
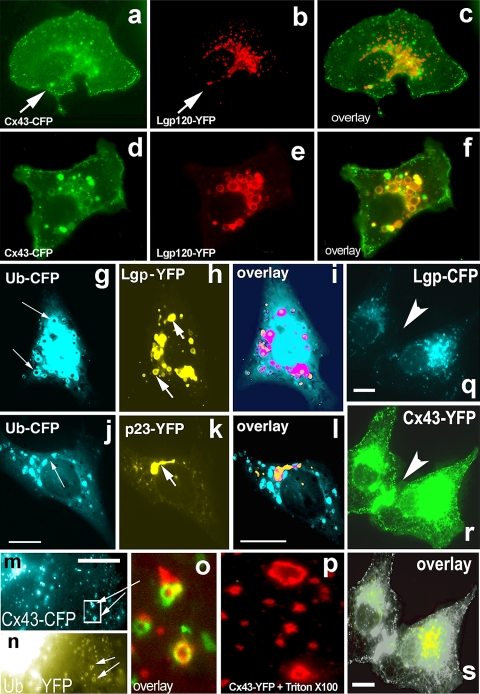
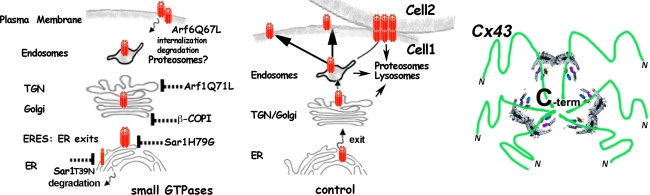
Connexins are four-transmembrane-domain proteins expressed in all vertebrates which form permeable gap junction channels that connect cells. Here, we analysed Connexin-43 (Cx43) transport to the plasma membrane and studied the effects of small GTPases acting along the secretory pathway. We show that both GTP- and GDP-restricted Sar1 prevents exit of Cx43 from the endoplasmic reticulum (ER), but only GTP-restricted Sar1 arrests Cx43 in COP II-coated ER exit sites and accumulates 14-3-3 proteins in the ER fraction. FRET-FLIM data confirm that already in ER exit sites Cx43 exists in oligomeric form, suggesting an in vivo role for 14-3-3 in Cx43 oligomerization. Exit of Cx43 from the ER can be blocked by other factors--such as expression of the beta subunit of the COP I coat or p50/dynamitin that acts on the microtubule-based dynein motor complex. GTP-restricted Arf1 blocks Cx43 in the Golgi. Lastly, we show that GTP-restricted Arf6 removes Cx43 gap junction plaques from the cell-cell interface and targets them to degradation. These data provide a molecular explanation of how small GTPases act to regulate Cx43 transport through the secretory pathway, facilitating or abolishing cell-cell communication through gap junctions.
Figures
Similar articles
-
Mechanisms of Cx43 and Cx26 transport to the plasma membrane and gap junction regeneration.J Cell Sci. 2005 Oct 1;118(Pt 19):4451-62. doi: 10.1242/jcs.02569. Epub 2005 Sep 13. J Cell Sci. 2005. PMID: 16159960
-
Defining a minimal motif required to prevent connexin oligomerization in the endoplasmic reticulum.J Biol Chem. 2005 Jun 3;280(22):21115-21. doi: 10.1074/jbc.M412612200. Epub 2005 Apr 7. J Biol Chem. 2005. PMID: 15817491
-
Identification of rab20 as a potential regulator of connexin 43 trafficking.Cell Commun Adhes. 2008 May;15(1):65-74. doi: 10.1080/15419060802014305. Cell Commun Adhes. 2008. PMID: 18649179
-
ER-to-Golgi transport: COP I and COP II function (Review).Mol Membr Biol. 2003 Jul-Sep;20(3):197-207. doi: 10.1080/0968768031000122548. Mol Membr Biol. 2003. PMID: 12893528 Review.
-
[Remodeling of cardiac gap junctions and arrhythmias].Sheng Li Xue Bao. 2011 Dec 25;63(6):586-92. Sheng Li Xue Bao. 2011. PMID: 22193455 Review. Chinese.
Cited by
-
Gap junctions.Compr Physiol. 2012 Jul;2(3):1981-2035. doi: 10.1002/cphy.c110051. Compr Physiol. 2012. PMID: 23723031 Free PMC article. Review.
-
Connexin 43 and CaV1.2 Ion Channel Trafficking in Healthy and Diseased Myocardium.Circ Arrhythm Electrophysiol. 2016 Jun;9(6):e001357. doi: 10.1161/CIRCEP.115.001357. Circ Arrhythm Electrophysiol. 2016. PMID: 27266274 Free PMC article. Review. No abstract available.
-
Progesterone Via its Type-A Receptor Promotes Myometrial Gap Junction Coupling.Sci Rep. 2017 Oct 17;7(1):13357. doi: 10.1038/s41598-017-13488-9. Sci Rep. 2017. PMID: 29042599 Free PMC article.
-
Fluorescence Guided Raman Spectroscopy enables the training of robust support vector machines for the detection of tumour marker proteins.Sci Rep. 2025 Jul 3;15(1):23711. doi: 10.1038/s41598-025-08425-0. Sci Rep. 2025. PMID: 40610529 Free PMC article.
-
Connexin43 hemichannel-mediated regulation of connexin43.PLoS One. 2013;8(2):e58057. doi: 10.1371/journal.pone.0058057. Epub 2013 Feb 27. PLoS One. 2013. PMID: 23460926 Free PMC article.
References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1083/jcb.131.4.875', 'is_inner': False, 'url': 'https://doi.org/10.1083/jcb.131.4.875'}, {'type': 'PMC', 'value': 'PMC2200014', 'is_inner': False, 'url': 'https://pmc.ncbi.nlm.nih.gov/articles/PMC2200014/'}, {'type': 'PubMed', 'value': '7490291', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/7490291/'}]}
- Aridor M, Bannykh SI, Rowe T, Balch WE (1995) Sequential coupling between COPII and COPI vesicle coats in endoplasmic reticulum to Golgi transport. J Cell Biol 131:875–893 - PMC - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/S0306-4522(01)00492-4', 'is_inner': False, 'url': 'https://doi.org/10.1016/s0306-4522(01)00492-4'}, {'type': 'PubMed', 'value': '11784696', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/11784696/'}]}
- Baxter HC, Liu WL, Forster JL, Aitken A, Fraser JR (2002) Immunolocalisation of 14-3-3 isoforms in normal and scrapie-infected murine brain. Neuroscience 109:5–14 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/S0092-8674(03)01079-1', 'is_inner': False, 'url': 'https://doi.org/10.1016/s0092-8674(03)01079-1'}, {'type': 'PubMed', 'value': '14744428', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/14744428/'}]}
- Bonifacino JS, Glick BS (2004) The mechanisms of vesicle budding and fusion. Cell 116:153–166 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '4473832', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/4473832/'}]}
- Bozhkova VP, Kvavilashvili IS, Rott NN, Chailakhyan LM (1974) Measurement of electrical coupling between cells of axolotl embryos during cleavage divisions. Sov J Dev Biol 4:480–482 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1083/jcb.139.2.469', 'is_inner': False, 'url': 'https://doi.org/10.1083/jcb.139.2.469'}, {'type': 'PMC', 'value': 'PMC2139801', 'is_inner': False, 'url': 'https://pmc.ncbi.nlm.nih.gov/articles/PMC2139801/'}, {'type': 'PubMed', 'value': '9334349', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/9334349/'}]}
- Burkhardt JK, Echeverri CJ, Nilsson T, Vallee RB (1997) Overexpression of the dynamitin (p50) subunit of the dynactin complex disrupts dynein-dependent maintenance of membrane organelle distribution. J Cell Biol 139:469–484 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
