

YA is needed for proper nuclear organization to transition between meiosis and mitosis in Drosophila
- PMID: 19627584
- PMCID: PMC2724486
- DOI: 10.1186/1471-213X-9-43
YA is needed for proper nuclear organization to transition between meiosis and mitosis in Drosophila
Abstract
Background: The Drosophila YA protein is required to initiate the embryonic cleavage divisions. After egg activation, YA enters nuclei and interacts with chromatin and the nuclear lamina. This study was designed to define more precisely the events prior to the first cleavage division that are dependent upon YA.
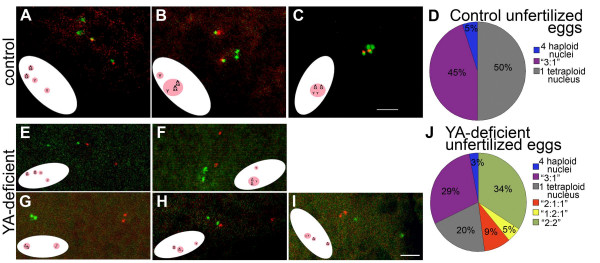
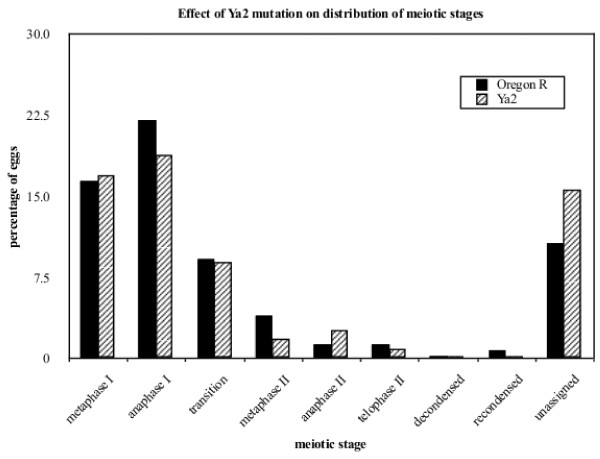
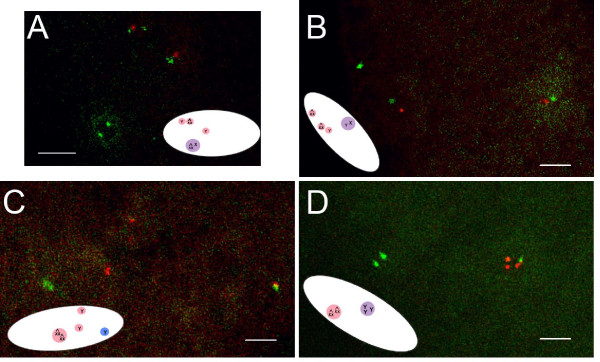
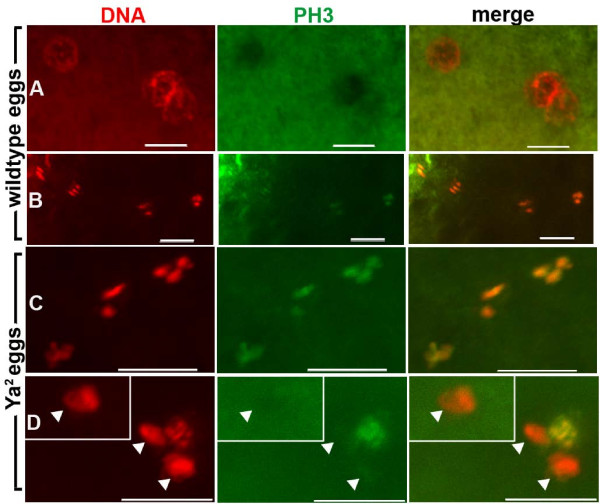
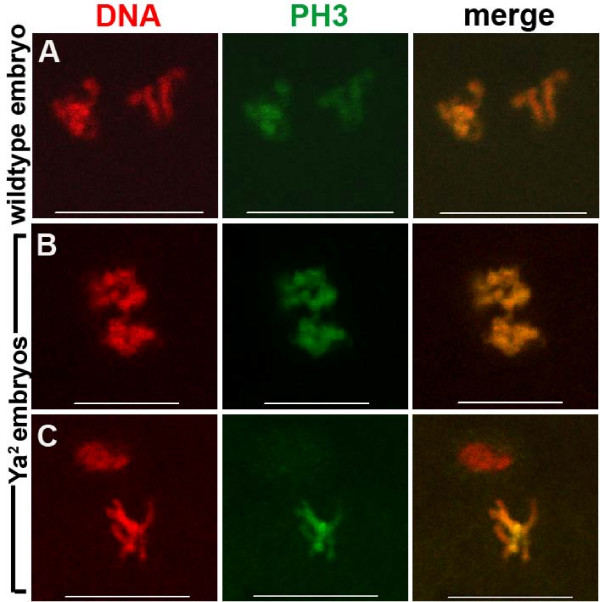
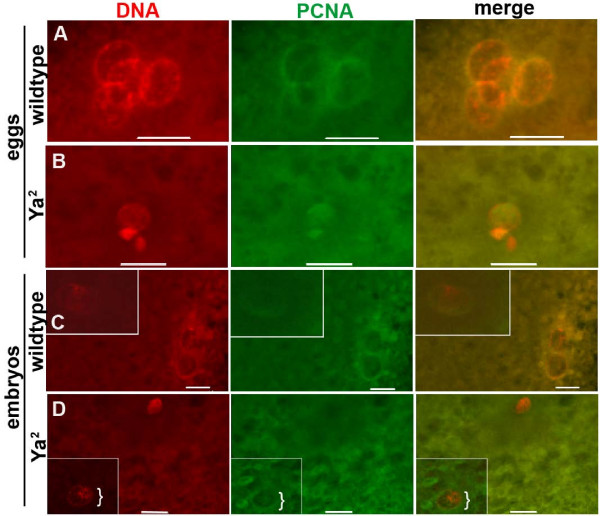
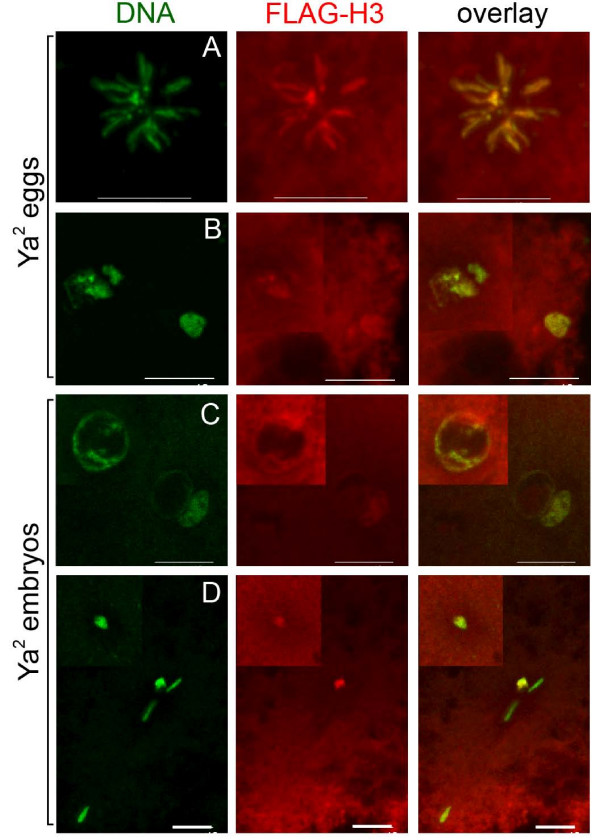
Results: We find that meiosis is completed normally in the absence of YA function. The first defects in embryos and eggs from mutant mothers first appear just after the completion of meiosis, and are seen as abnormal associations among the resultant haploid nuclei. These defects are associated with asynchronies in the cell cycle-dependent chromatin condensation state of the haploid nuclei. However, we find evidence of DNA replication in the absence of YA function.
Conclusion: Our data suggest YA function is needed at a control point, following meiosis II and the initiation of the first postmeiotic S phase, which is sensitive to the chromatin condensation state of the haploid meiotic products.
Figures
Similar articles
-
The Drosophila nuclear lamina protein YA binds to DNA and histone H2B with four domains.Mol Biol Cell. 2002 Feb;13(2):558-69. doi: 10.1091/mbc.01-07-0336. Mol Biol Cell. 2002. PMID: 11854412 Free PMC article.
-
Mutational analyses of fs(1)Ya, an essential, developmentally regulated, nuclear envelope protein in Drosophila.Genetics. 1995 Dec;141(4):1473-81. doi: 10.1093/genetics/141.4.1473. Genetics. 1995. PMID: 8601487 Free PMC article.
-
Mutations in the chromosomal passenger complex and the condensin complex differentially affect synaptonemal complex disassembly and metaphase I configuration in Drosophila female meiosis.Genetics. 2009 Mar;181(3):875-87. doi: 10.1534/genetics.108.097741. Epub 2008 Dec 22. Genetics. 2009. PMID: 19104074 Free PMC article.
-
Cohesin in determining chromosome architecture.Exp Cell Res. 2012 Jul 15;318(12):1386-93. doi: 10.1016/j.yexcr.2012.03.016. Epub 2012 Mar 24. Exp Cell Res. 2012. PMID: 22472347 Review.
-
Polo kinase--meiotic cell cycle coordinator.Cell Cycle. 2003 Sep-Oct;2(5):400-2. Cell Cycle. 2003. PMID: 12963824 Review. No abstract available.
Cited by
-
Developmental control of oocyte maturation and egg activation in metazoan models.Cold Spring Harb Perspect Biol. 2011 Oct 1;3(10):a005553. doi: 10.1101/cshperspect.a005553. Cold Spring Harb Perspect Biol. 2011. PMID: 21709181 Free PMC article. Review.
-
Phospho-regulation pathways during egg activation in Drosophila melanogaster.Genetics. 2013 Sep;195(1):171-80. doi: 10.1534/genetics.113.150110. Epub 2013 Jun 21. Genetics. 2013. PMID: 23792954 Free PMC article.
-
Protein phosphorylation changes reveal new candidates in the regulation of egg activation and early embryogenesis in D. melanogaster.Dev Biol. 2012 Oct 1;370(1):125-34. doi: 10.1016/j.ydbio.2012.07.024. Epub 2012 Jul 31. Dev Biol. 2012. PMID: 22884528 Free PMC article.
-
The Drosophila nucleoporin ELYS is required for parental chromosome arrangement at fertilization.G3 (Bethesda). 2025 Jul 9;15(7):jkaf104. doi: 10.1093/g3journal/jkaf104. G3 (Bethesda). 2025. PMID: 40359232 Free PMC article.
-
Genome-wide analysis of mRNA decay patterns during early Drosophila development.Genome Biol. 2010;11(9):R93. doi: 10.1186/gb-2010-11-9-r93. Epub 2010 Sep 21. Genome Biol. 2010. PMID: 20858238 Free PMC article.
References
-
- Heifetz Y, Yu J, Wolfner MF. Ovulation triggers activation of Drosophila oocytes. Dev Biol. 2001;234:416–24. - PubMed
-
- Mahowald AP, Goralski TJ, Caulton JH. In vitro activation of Drosophila eggs. Dev Biol. 1983;98:437–45. - PubMed
-
- Page AW, Orr-Weaver TL. Activation of the meiotic divisions in Drosophila oocytes. Dev Biol. 1997;183:195–207. - PubMed
-
- Horner VL, Czank A, Jang JK, Singh N, Williams BC, Puro J, Kubli E, Hanes SD, McKim KS, Wolfner MF, Goldberg ML. The Drosophila calcipressin sarah is required for several aspects of egg activation. Curr Biol. 2006;16:1441–6. - PubMed
-
- Takeo S, Tsuda M, Akahori S, Matsuo T, Aigaki T. The calcineurin regulator sra plays an essential role in female meiosis in Drosophila. Curr Biol. 2006;16:1435–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
