

Molecular phylogenetics and evolutionary history of ariid catfishes revisited: a comprehensive sampling
- PMID: 19627593
- PMCID: PMC2726142
- DOI: 10.1186/1471-2148-9-175
Molecular phylogenetics and evolutionary history of ariid catfishes revisited: a comprehensive sampling
Abstract
Background: Ariids or sea catfishes are one of the two otophysan fish families (out of about 67 families in four orders) that inhabit mainly marine and brackish waters (although some species occur strictly in fresh waters). The group includes over 150 species placed in approximately 29 genera and two subfamilies (Galeichthyinae and Ariinae). Despite their global distribution, ariids are largely restricted to the continental shelves due in part to their specialized reproductive behavior (i.e., oral incubation). Thus, among marine fishes, ariids offer an excellent opportunity for inferring historical biogeographic scenarios. Phylogenetic hypotheses available for ariids have focused on restricted geographic areas and comprehensive phylogenies are still missing. This study inferred phylogenetic hypotheses for 123 ariid species in 28 genera from different biogeographic provinces using both mitochondrial and nuclear sequences (up to approximately 4 kb).
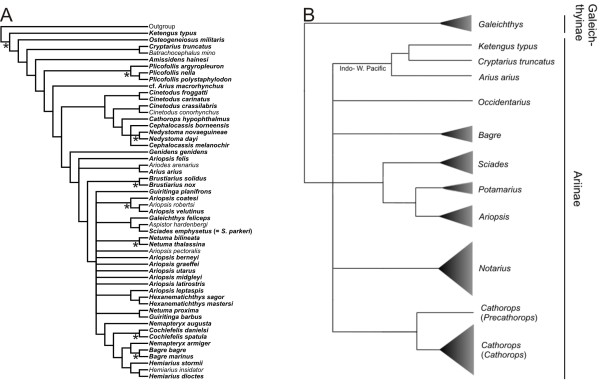
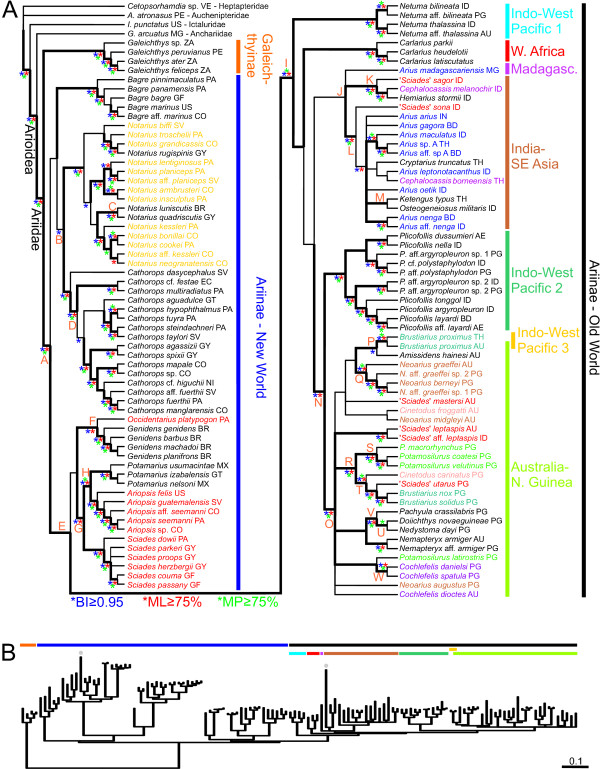
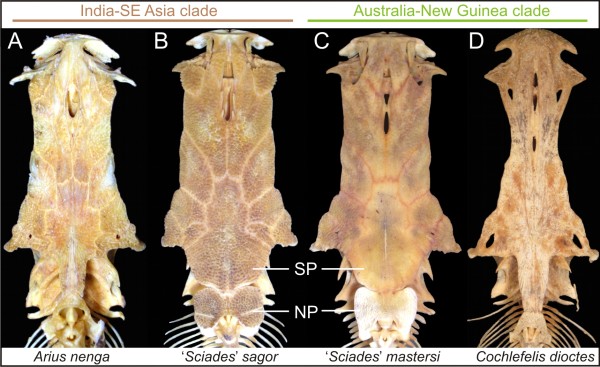
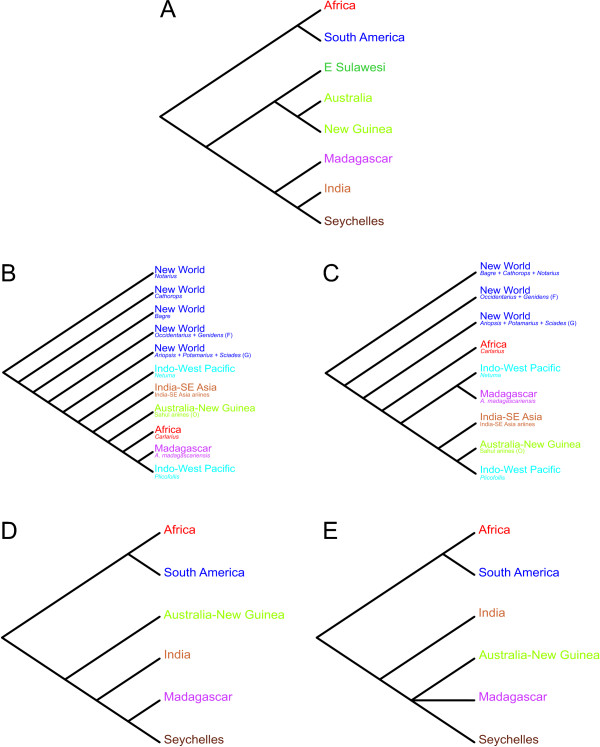
Results: While the topologies obtained support the monophyly of basal groups, up to ten genera validated in previous morphological studies were incongruent with the molecular topologies. New World ariines were recovered as paraphyletic and Old World ariines were grouped into a well-supported clade that was further divided into subclades mainly restricted to major Gondwanan landmasses. A general area cladogram derived from the area cladograms of ariines and three other fish groups was largely congruent with the geological area cladogram of Gondwana. Nonetheless, molecular clock estimations provided variable results on the timing of ariine diversification (approximately 105-41 mya).
Conclusion: This study provides the most comprehensive phylogeny of sea catfishes to date and highlights the need for re-assessment of their classification. While from a topological standpoint the evolutionary history of ariines is mostly congruent with vicariance associated with the sequence of events during Gondwanan fragmentation, ambiguous divergence time estimations hinders assessing the vicariant hypothesis on a temporal framework. Further examination of ariid fossils might provide the basis for more accurate inferences on the timing of ariine diversification.
Figures
Similar articles
-
Systematics and biogeography of New World sea catfishes (Siluriformes: Ariidae) as inferred from mitochondrial, nuclear, and morphological evidence.Mol Phylogenet Evol. 2007 Oct;45(1):339-57. doi: 10.1016/j.ympev.2007.02.022. Epub 2007 Feb 28. Mol Phylogenet Evol. 2007. PMID: 17475516
-
Molecular phylogenetics supports multiple evolutionary transitions from marine to freshwater habitats in ariid catfishes.Mol Phylogenet Evol. 2010 Apr;55(1):249-258. doi: 10.1016/j.ympev.2009.12.018. Epub 2010 Jan 4. Mol Phylogenet Evol. 2010. PMID: 20045737
-
Molecular clocks provide new insights into the evolutionary history of Galeichthyine sea catfishes.Evolution. 2009 May;63(5):1232-43. doi: 10.1111/j.1558-5646.2009.00640.x. Epub 2008 Feb 2. Evolution. 2009. PMID: 19187251
-
Molecular phylogeny of grunts (Teleostei, Haemulidae), with an emphasis on the ecology, evolution, and speciation history of new world species.BMC Evol Biol. 2012 Apr 26;12:57. doi: 10.1186/1471-2148-12-57. BMC Evol Biol. 2012. PMID: 22537107 Free PMC article.
-
Evolutionary history of Otophysi (Teleostei), a major clade of the modern freshwater fishes: Pangaean origin and Mesozoic radiation.BMC Evol Biol. 2011 Jun 22;11:177. doi: 10.1186/1471-2148-11-177. BMC Evol Biol. 2011. PMID: 21693066 Free PMC article.
Cited by
-
Bayesian Divergence-Time Estimation with Genome-Wide Single-Nucleotide Polymorphism Data of Sea Catfishes (Ariidae) Supports Miocene Closure of the Panamanian Isthmus.Syst Biol. 2018 Jul 1;67(4):681-699. doi: 10.1093/sysbio/syy006. Syst Biol. 2018. PMID: 29385552 Free PMC article.
-
Evolution of opercle bone shape along a macrohabitat gradient: species identification using mtDNA and geometric morphometric analyses in neotropical sea catfishes (Ariidae).Ecol Evol. 2016 Jul 22;6(16):5817-30. doi: 10.1002/ece3.2334. eCollection 2016 Aug. Ecol Evol. 2016. PMID: 27547357 Free PMC article.
-
New phylogenetic insights into the African catfish families Mochokidae and Austroglanididae.J Fish Biol. 2022 May;100(5):1171-1186. doi: 10.1111/jfb.15014. Epub 2022 Mar 9. J Fish Biol. 2022. PMID: 35184288 Free PMC article.
-
Marine catfishes (Ariidae-Siluriformes) from the Coastal Amazon: mitochondrial DNA barcode for a recent diversification group?PeerJ. 2024 Aug 28;12:e17581. doi: 10.7717/peerj.17581. eCollection 2024. PeerJ. 2024. PMID: 39221281 Free PMC article.
-
Morphological and molecular (28S rRNA) data of monogeneans (Platyhelminthes) infecting the gill lamellae of marine fishes in the Campeche Bank, southwest Gulf of Mexico.Zookeys. 2018 Sep 11;(783):125-161. doi: 10.3897/zookeys.783.26218. eCollection 2018. Zookeys. 2018. PMID: 30275730 Free PMC article.
References
-
- Grande L, Eastman JT. A Review of Antarctic ichthyofaunas in the light of new fossil discoveries. Paleontology. 1986;29:113–137.
-
- All Catfish Species Inventory http://silurus.acnatsci.org/
-
- Ferraris CJ. Checklist of catfishes, recent and fossil (Osteichthyes:Siluriformes), and catalogue of siluriform primary types. Zootaxa. 2007. p. 548.
-
- Mo T. Anatomy, Relationships and Systematics of the Bagridae (Teleostei: Siluroidei) with a Hypothesis of Siluroid Phylogeny (Theses Zoologicae) Vol. 17. Koenigstein: Koeltz; 1991.
-
- de Pinna MCC. Phylogenetic relationships of Neotropical Siluriformes (Teleostei: Ostariophysi): historical overview and synthesis of hypotheses. In: Malabarba LR, Reis RE, Vari RP, Lucena ZMS, Lucena CAS, editor. Phylogeny and Classification of Neotropical Fishes. Porto Alegre: EDIPUCRS; 1998.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
