

Transcriptional profiling of maturing tomato (Solanum lycopersicum L.) microspores reveals the involvement of heat shock proteins, ROS scavengers, hormones, and sugars in the heat stress response
- PMID: 19628571
- PMCID: PMC2736902
- DOI: 10.1093/jxb/erp234
Transcriptional profiling of maturing tomato (Solanum lycopersicum L.) microspores reveals the involvement of heat shock proteins, ROS scavengers, hormones, and sugars in the heat stress response
Abstract
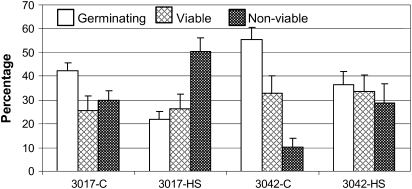
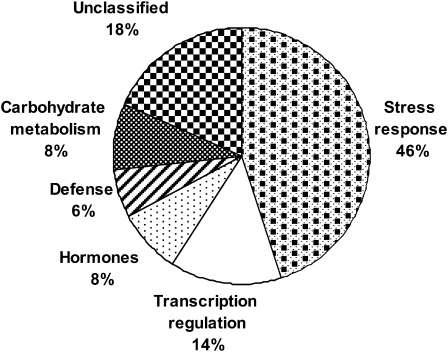
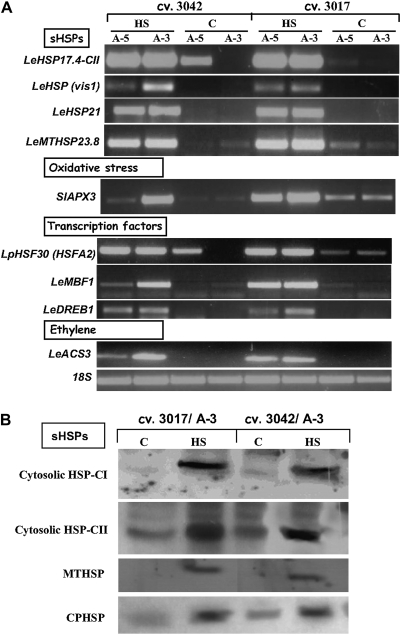
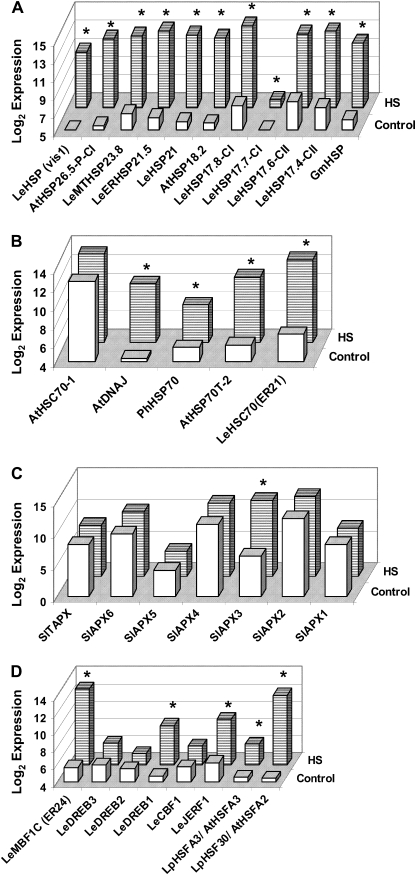
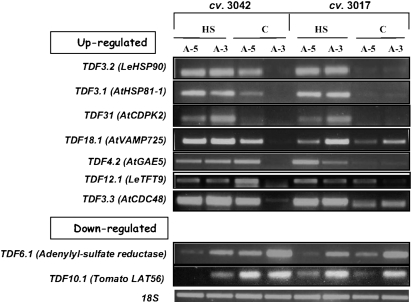
Above-optimal temperatures reduce yield in tomato largely because of the high heat stress (HS) sensitivity of the developing pollen grains. The high temperature response, especially at this most HS-sensitive stage of the plant, is poorly understood. To obtain an overview of molecular mechanisms underlying the HS response (HSR) of microspores, a detailed transcriptomic analysis of heat-stressed maturing tomato microspores was carried out using a combination of Affymetrix Tomato Genome Array and cDNA-amplified fragment length polymorphism (AFLP) techniques. The results were corroborated by reverse transcription-PCR (RT-PCR) and immunoblot analyses. The data obtained reveal the involvement of specific members of the small heat shock protein (HSP) gene family, HSP70 and HSP90, in addition to the HS transcription factors A2 (HSFA2) and HSFA3, as well as factors other than the classical HS-responsive genes. The results also indicate HS regulation of reactive oxygen species (ROS) scavengers, sugars, plant hormones, and regulatory genes that were previously implicated in other types of stress. The use of cDNA-AFLP enabled the detection of genes representing pollen-specific functions that are missing from the tomato Affymetrix chip, such as those involved in vesicle-mediated transport and a pollen-specific, calcium-dependent protein kinase (CDPK2). For several genes, including LeHSFA2, LeHSP17.4-CII, as well as homologues of LeHSP90 and AtVAMP725, higher basal expression levels were detected in microspores of cv. Hazera 3042 (a heat-tolerant cultivar) compared with microspores of cv. Hazera 3017 (a heat-sensitive cultivar), marking these genes as candidates for taking part in microspore thermotolerance. This work provides a comprehensive analysis of the molecular events underlying the HSR of maturing microspores of a crop plant, tomato.
Figures
References
-
- Aloni B, Peet M, Pharr M, Karni L. The effect of high temperature and high atmospheric CO2 on carbohydrate changes in bell pepper (Capsicum annuum) pollen in relation to its germination. Physiologia Plantarum. 2001;112:505–512. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. Journal of Molecular Biology. 1990;215:403–410. - PubMed
-
- Bao YM, Wang JF, Huang J, Zhang HS. Cloning and characterization of three genes encoding Qb-SNARE proteins in rice. Molecular Genetics and Genomics. 2008;279:291–301. - PubMed
-
- Baniwal SK, Bharti K, Chan KY, et al. Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. Journal of Biosciences. 2004;29:471–487. - PubMed
