

Taming the tiger by the tail: modulation of DNA damage responses by telomeres
- PMID: 19629039
- PMCID: PMC2722249
- DOI: 10.1038/emboj.2009.176
Taming the tiger by the tail: modulation of DNA damage responses by telomeres
Abstract
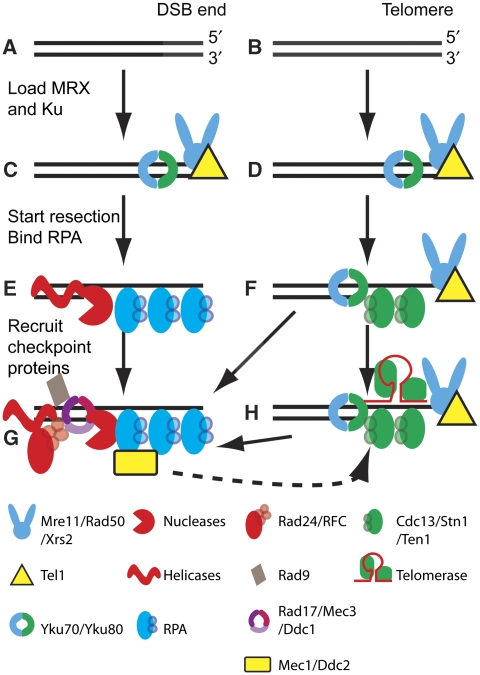
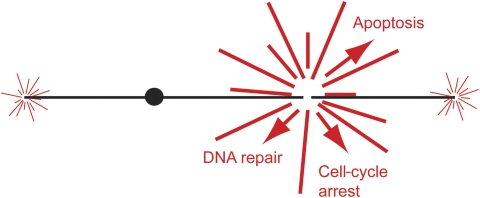
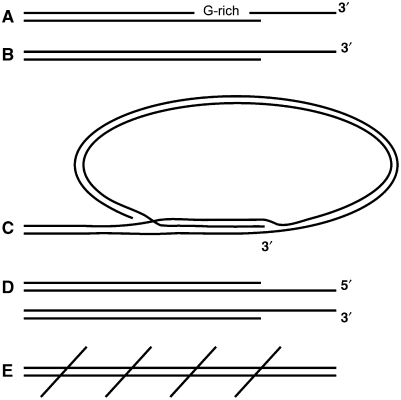
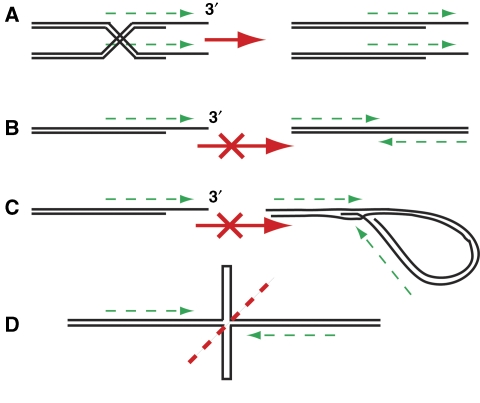
Telomeres are by definition stable and inert chromosome ends, whereas internal chromosome breaks are potent stimulators of the DNA damage response (DDR). Telomeres do not, as might be expected, exclude DDR proteins from chromosome ends but instead engage with many DDR proteins. However, the most powerful DDRs, those that might induce chromosome fusion or cell-cycle arrest, are inhibited at telomeres. In budding yeast, many DDR proteins that accumulate most rapidly at double strand breaks (DSBs), have important functions in physiological telomere maintenance, whereas DDR proteins that arrive later tend to have less important functions. Considerable diversity in telomere structure has evolved in different organisms and, perhaps reflecting this diversity, different DDR proteins seem to have distinct roles in telomere physiology in different organisms. Drawing principally on studies in simple model organisms such as budding yeast, in which many fundamental aspects of the DDR and telomere biology have been established; current views on how telomeres harness aspects of DDR pathways to maintain telomere stability and permit cell-cycle division are discussed.
Figures
References
-
- Addinall SG, Downey M, Yu M, Zubko MK, Dewar J, Leake A, Hallinan J, Shaw O, James K, Wilkinson DJ, Wipat A, Durocher D, Lydall D (2008) A genomewide suppressor and enhancer analysis of cdc13-1 reveals varied cellular processes influencing telomere capping in Saccharomyces cerevisiae. Genetics 180: 2251–2266 - PMC - PubMed
-
- Ahmed S, Hodgkin J (2000) MRT-2 checkpoint protein is required for germline immortality and telomere replication in C. elegans [see comments]. Nature 403: 159–164 - PubMed
-
- Artandi SE, Attardi LD (2005) Pathways connecting telomeres and p53 in senescence, apoptosis, and cancer. Biochem Biophys Res Commun 331: 881–890 - PubMed
-
- Aubert G, Lansdorp PM (2008) Telomeres and aging. Physiol Rev 88: 557–579 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
