

Review
doi: 10.1089/hum.2009.135.
The case of oncolytic viruses versus the immune system: waiting on the judgment of Solomon
Affiliations
- PMID: 19630549
- PMCID: PMC2829276
- DOI: 10.1089/hum.2009.135
Item in Clipboard
Review
The case of oncolytic viruses versus the immune system: waiting on the judgment of Solomon
Hum Gene Ther.
2009 Oct.
Abstract
The three-way interaction between oncolytic viruses, the tumor microenvironment, and the immune system is critical to the outcome of antitumor therapy. Classically, the immune system is thought to limit the efficacy of therapy, leading to viral clearance. However, preclinical and clinical data suggest that in some cases virotherapy may in fact act as cancer immunotherapy. In this review we discuss the ability of oncolytic viruses to alter the immunogenic milieu of the tumor microenvironment, and the role of innate and adaptive immunity in both restricting and augmenting therapy. Strategies to improve virotherapy by immunomodulation, including suppression or enhancement of the innate and adaptive responses, are discussed.
Figures
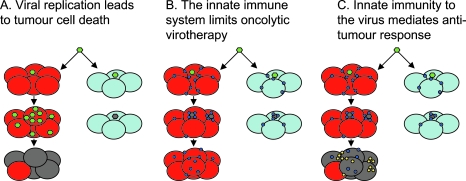
Different views on the mechanisms of oncolytic virotherapy. (A) Therapy from viral replication, spread, and lysis. Oncolytic virus (green) can infect either tumor (red) or normal (light blue) cells. The virus is either engineered to replicate only in tumor cells, or exploits defects in the antiviral innate immune response of tumor cells that allows its progressive spread through tumor but not normal cells. The antitumor effect relies solely on viral replication (dying tumor cells are gray). (B) Innate immunity to the virus is the enemy of oncolytic virotherapy. Viral infection of a tumor must also take into account the infiltrating immune cells that sense and respond to viral infection (blue circles). On exposure to virus, these cells secrete potent antiviral cytokines/interferons and other effector molecules. Many tumors are not completely defective in their responses to type I interferons and this innate activation within the tumor environment prevents viral replication and spread—even in tumor cells. This leads to clearance of the virus, extinction of the infection, and vastly reduced oncolysis and therapy. (C) Innate immunity to the virus is oncolytic virotherapy. In this view, the antiviral cytokines and interferons released by tumor-infiltrating immune cells in response to virus have significant antitumor activity themselves. These molecules act to shut down viral replication but also have directly antitumor effects by (1) killing tumor cells directly and/or (2) recruiting further antitumor immune effectors into the tumor (yellow circles), which finish off the job themselves. In this model, antitumor efficacy is mediated by the bystander killing of tumor cells induced by the immune response to viral infection.
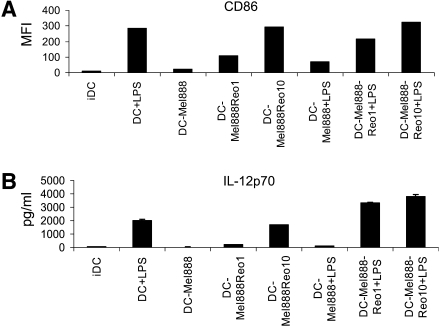
Reovirus activates tumor-loaded DCs, reversing their hyporesponsiveness to LPS. DCs were loaded with Mel888 cells (at a 1:3 ratio) ± reovirus (Reolysin) overnight, ±LPS (250 ng/ml) added after 8 hr. (A) After 24 hr CD86 expression on DCs was determined by flow cytometry. Median fluorescence intensity (MFI) is shown. IDC, immature dendritic cells. (B) IL-12p70 secretion was determined by ELISA of 24-hr supernatants. Error bars indicate the SE.
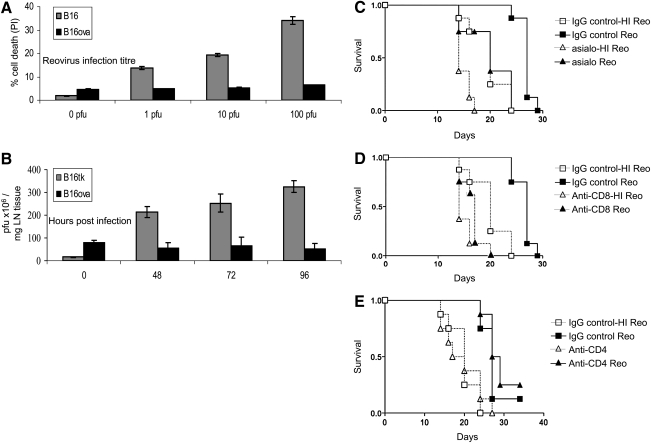
The B16ova cell line is resistant in vitro to reovirus, but regresses in vivo in an NK cell- and CD8+ T cell-dependent manner. (A) B16 and B16ova cell lines were infected with reovirus (Reolysin) for 48 hr, before assessment of cell death by administration of propidium iodide (PI). Error bars, SE. (B) C57BL/6 mice were seeded subcutaneously with 5 × 105 B16 or B16ova cells. After 7 days tumors were injected with a single dose of 5 × 108 plaque-forming units (PFU) of reovirus. Tumors were harvested at the indicated time points, and viral titer was determined by plaque assay. Error bars, SE. LN, lymph node. (C–E) C57BL/6 mice were seeded subcutaneously with 5 × 105 B16ova cells. A 0.1-mg amount of depleting antibody [(C) anti-NK (asialo), (D) anti-CD8, (E) anti-CD4] or control antibody was given intraperitoneally 4 days after tumor implantation (n = 8 per group), and every week for the duration of the experiment. Reovirus (5 × 108 PFU) or heat-inactivated reovirus, as a control, was injected intratumorally on days 7, 9, 11, and 13 after tumor implantation. (C) IgG control + reovirus versus NK cell depletion + reovirus; p = 0.008. (D) IgG control + reovirus versus CD8+ cell depletion + reovirus; p < 0.0001. (E) IgG control + reovirus versus CD4+ cell depletion + reovirus; p = 0.28 (by log-rank test).
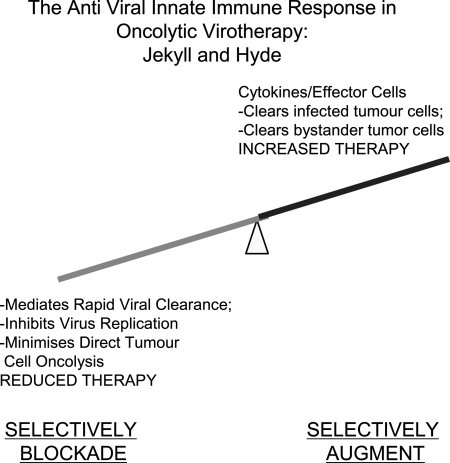
Future perspectives. The immune response to oncolytic virus comprises multiple components, some positive and some detrimental to therapy. In the future, it may be possible to identify those components that are specifically responsible for mediating viral clearance (detrimental to therapy) and those that are responsible for mediating antitumor efficacy (positive effect on therapy). Furthermore, it may be possible to block the former (such as the antiviral IFN-α/β response) while simultaneously augmenting the latter (expression of potently antitumor cytokines and interferons that are induced by IFN-α/β but that, per se, have no antiviral effects). In this way, virus design may be able to incorporate the best of both worlds—factors to reduce viral clearance (more direct oncolysis) and to increase recruitment of antitumor effectors (e.g., NK cell-recruiting cytokines).
References
-
- Aghi M. Martuza R.L. Oncolytic viral therapies: The clinical experience. Oncogene. 2005;24:7802–7816. - PubMed
-
- Albertsson P.A. Basse P.H. Hokland M. Goldfarb R.H. Nagelkerke J.F. Nannmark U. Kuppen P.J. NK cells and the tumour microenvironment: Implications for NK-cell function and anti-tumour activity. Trends Immunol. 2003;24:603–609. - PubMed
-
- Almand B. Resser J.R. Lindman B. Nadaf S. Clark J.I. Kwon E.D. Carbone D.P. Gabrilovich D.I. Clinical significance of defective dendritic cell differentiation in cancer. Clin. Cancer Res. 2000;6:1755–1766. - PubMed
-
- Andoniou C.E. Van Dommelen S.L. Voigt V. Andrews D.M. Brizard G. Asselin-Paturel C. Delale T. Stacey K.J. Trinchieri G. Degli-Esposti M.A. Interaction between conventional dendritic cells and natural killer cells is integral to the activation of effective antiviral immunity. Nat. Immunol. 2005;6:1011–1019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
