

Loss of Dnmt1 catalytic activity reveals multiple roles for DNA methylation during pancreas development and regeneration
- PMID: 19631206
- PMCID: PMC2759669
- DOI: 10.1016/j.ydbio.2009.07.017
Loss of Dnmt1 catalytic activity reveals multiple roles for DNA methylation during pancreas development and regeneration
Abstract
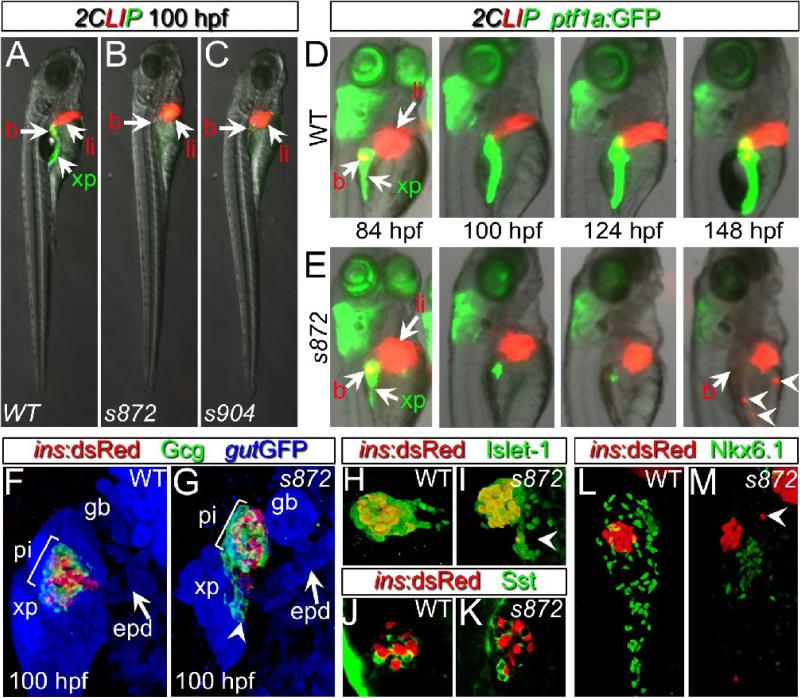
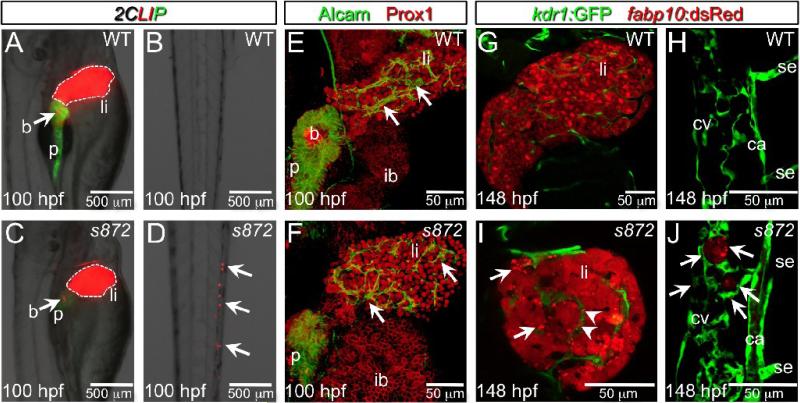
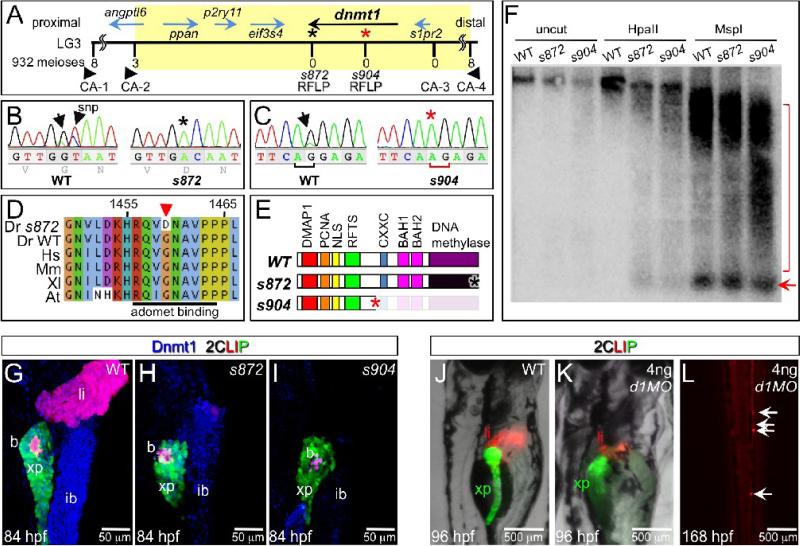
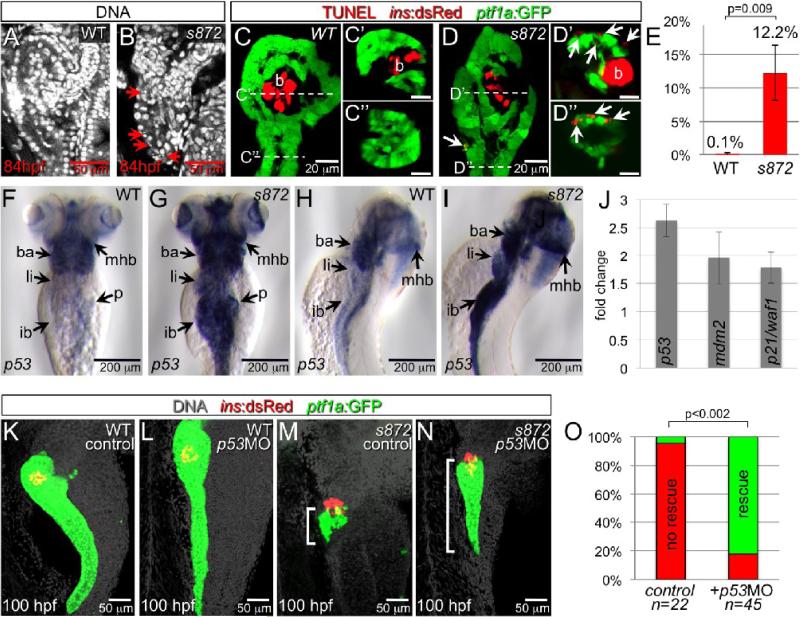
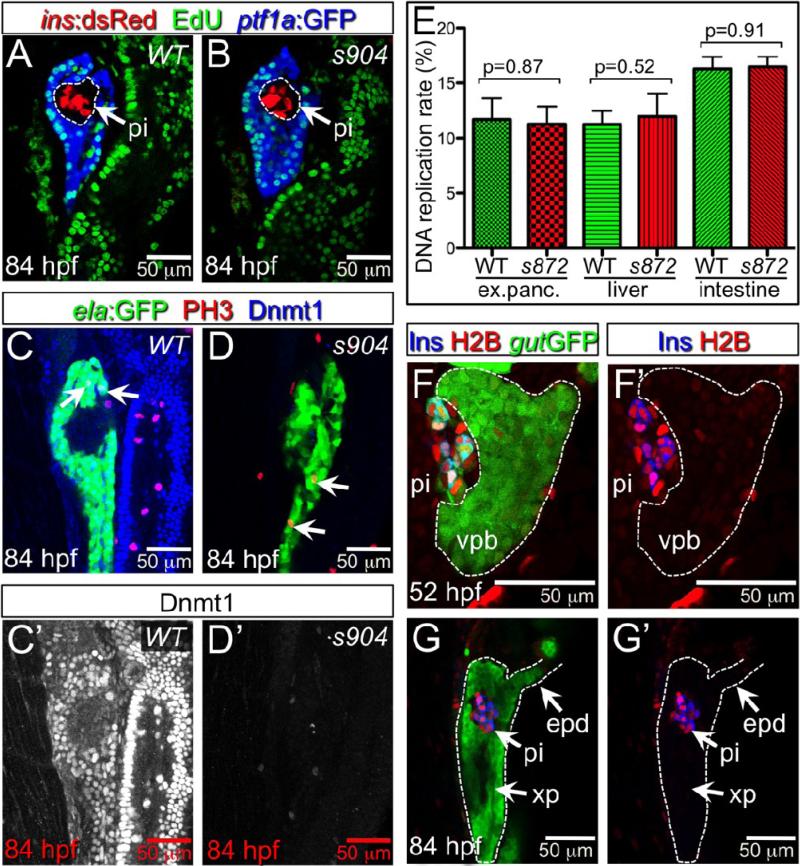
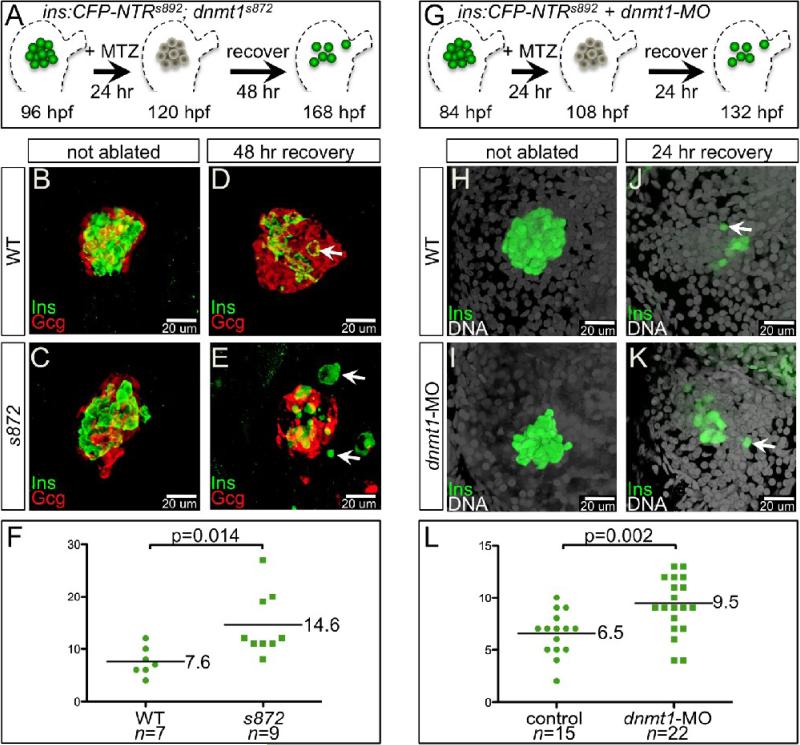
Developmental mechanisms regulating gene expression and the stable acquisition of cell fate direct cytodifferentiation during organogenesis. Moreover, it is likely that such mechanisms could be exploited to repair or regenerate damaged organs. DNA methyltransferases (Dnmts) are enzymes critical for epigenetic regulation, and are used in concert with histone methylation and acetylation to regulate gene expression and maintain genomic integrity and chromosome structure. We carried out two forward genetic screens for regulators of endodermal organ development. In the first, we screened for altered morphology of developing digestive organs, while in the second we screed for the lack of terminally differentiated cell types in the pancreas and liver. From these screens, we identified two mutant alleles of zebrafish dnmt1. Both lesions are predicted to eliminate dnmt1 function; one is a missense mutation in the catalytic domain and the other is a nonsense mutation that eliminates the catalytic domain. In zebrafish dnmt1 mutants, the pancreas and liver form normally, but begin to degenerate after 84 h post fertilization (hpf). Acinar cells are nearly abolished through apoptosis by 100 hpf, though neither DNA replication, nor entry into mitosis is halted in the absence of detectable Dnmt1. However, endocrine cells and ducts are largely spared. Surprisingly, dnmt1 mutants and dnmt1 morpholino-injected larvae show increased capacity for pancreatic beta cell regeneration in an inducible model of pancreatic beta cell ablation. Thus, our data suggest that Dnmt1 is dispensable for pancreatic duct or endocrine cell formation, but not for acinar cell survival. In addition, Dnmt1 may influence the differentiation of pancreatic beta cell progenitors or the reprogramming of cells toward the pancreatic beta cell fate.
Figures
References
-
- Beis D, Bartman T, Jin SW, Scott IC, D'Amico LA, Ober EA, Verkade H, Frantsve J, Field HA, Wehman A, Baier H, Tallafuss A, Bally-Cuif L, Chen JN, Stainier DY, Jungblut B. Genetic and cellular analyses of zebrafish atrioventricular cushion and valve development. Development. 2005;132:4193–204. - PubMed
-
- Biemar F, Argenton F, Schmidtke R, Epperlein S, Peers B, Driever W. Pancreas development in zebrafish: early dispersed appearance of endocrine hormone expressing cells and their convergence to form the definitive islet. Dev Biol. 2001;230:189–203. - PubMed
-
- Bird AP, Wolffe AP. Methylation-induced repression--belts, braces, and chromatin. Cell. 1999;99:451–4. - PubMed
-
- Bostick M, Kim JK, Esteve PO, Clark A, Pradhan S, Jacobsen SE. UHRF1 plays a role in maintaining DNA methylation in mammalian cells. Science. 2007;317:1760–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
