

Neurokinins inhibit low threshold inactivating K+ currents in capsaicin responsive DRG neurons
- PMID: 19631644
- PMCID: PMC2775709
- DOI: 10.1016/j.expneurol.2009.07.016
Neurokinins inhibit low threshold inactivating K+ currents in capsaicin responsive DRG neurons
Abstract
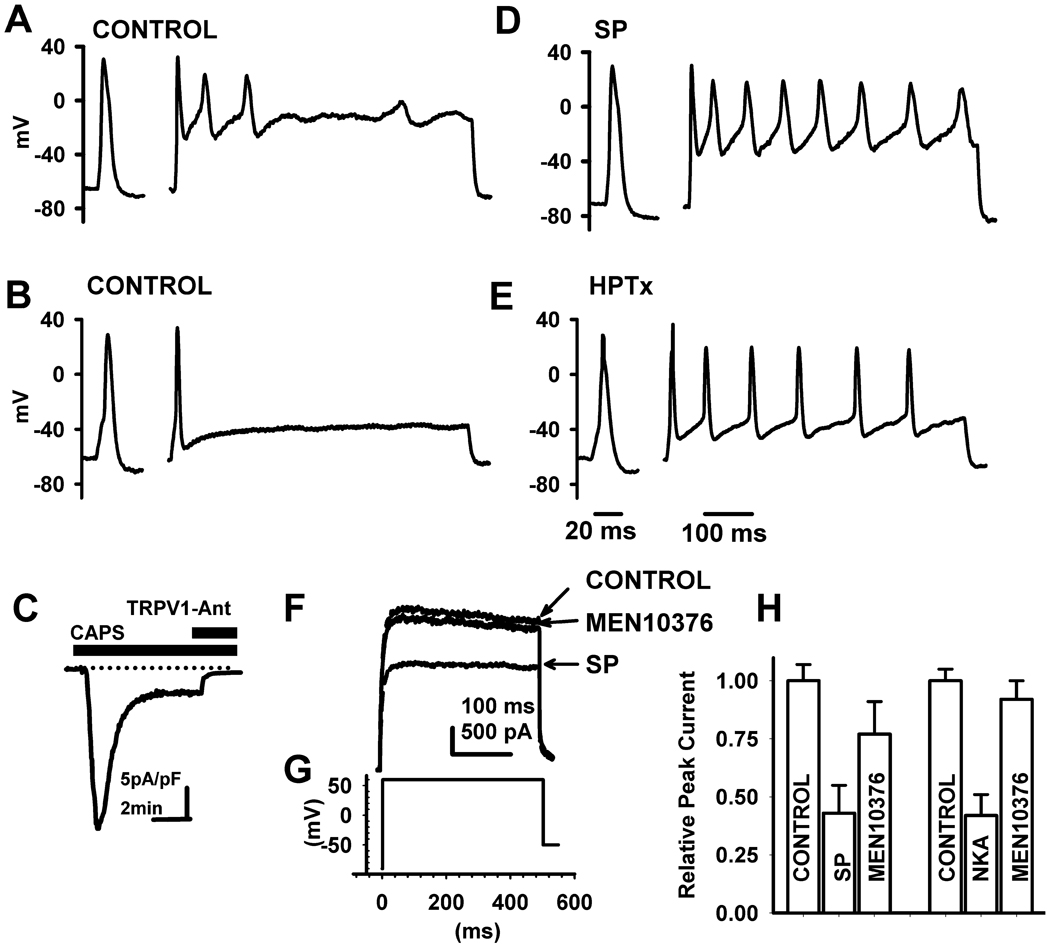
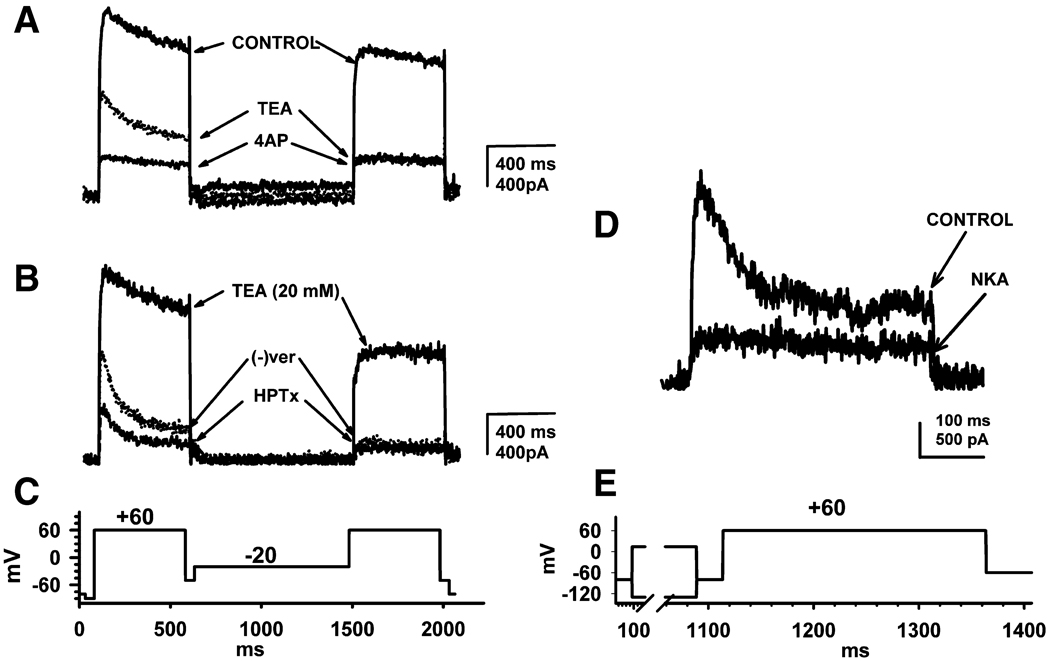
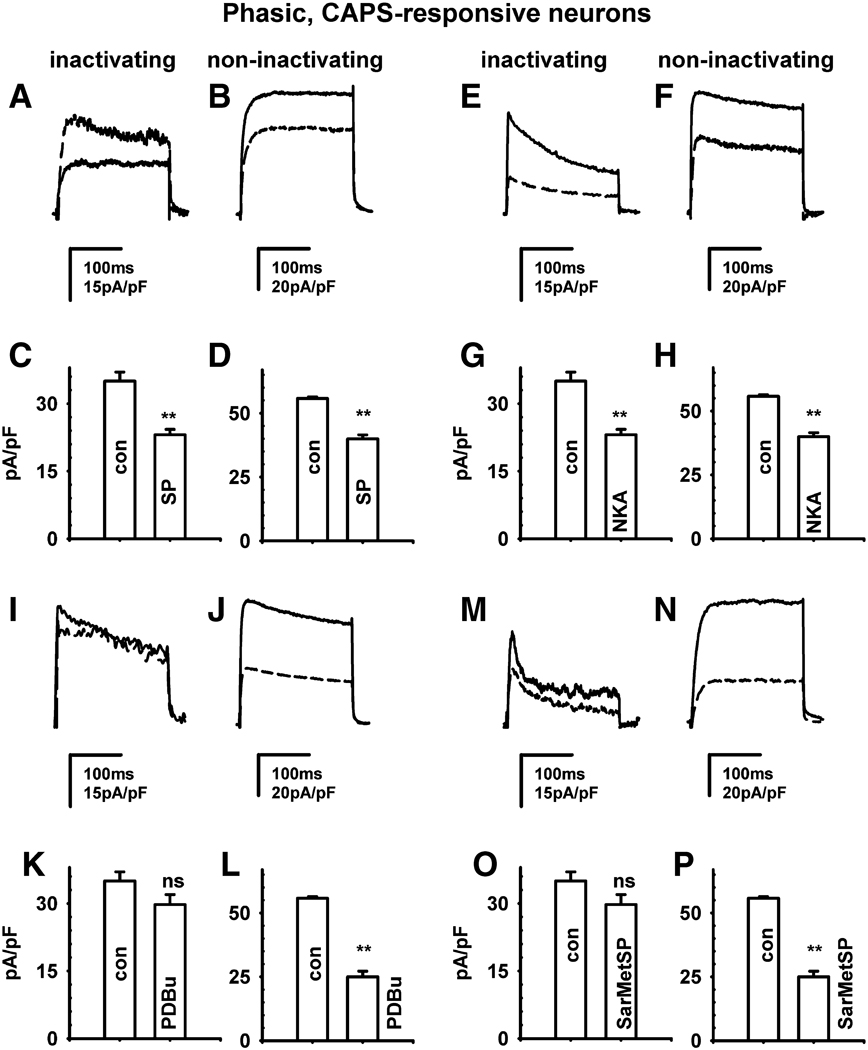
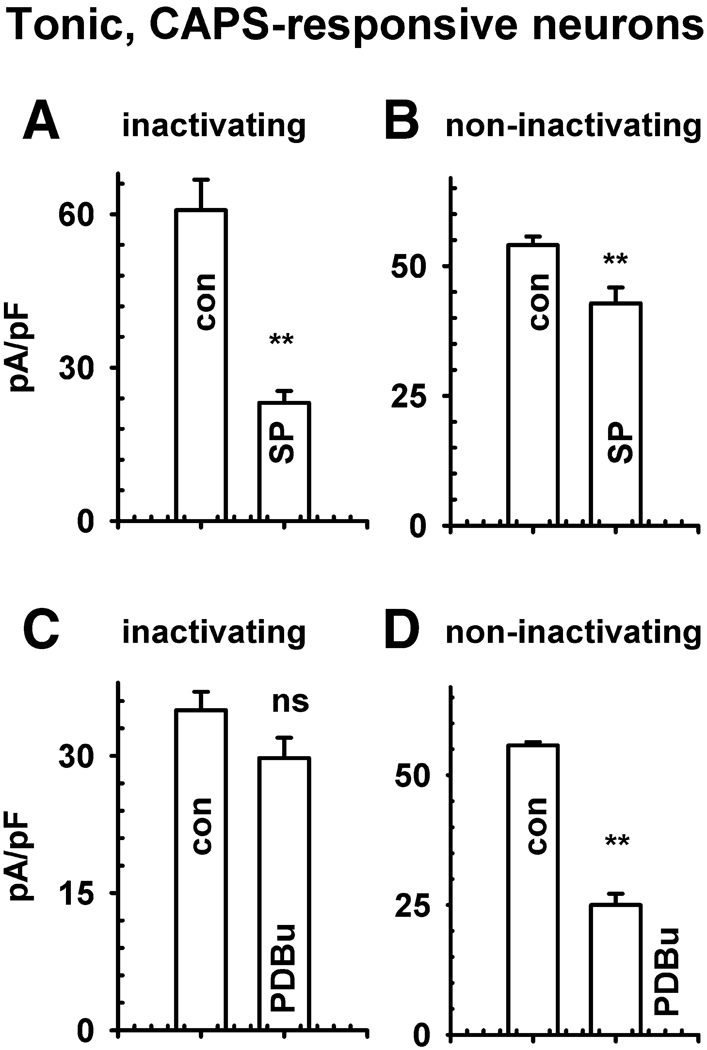
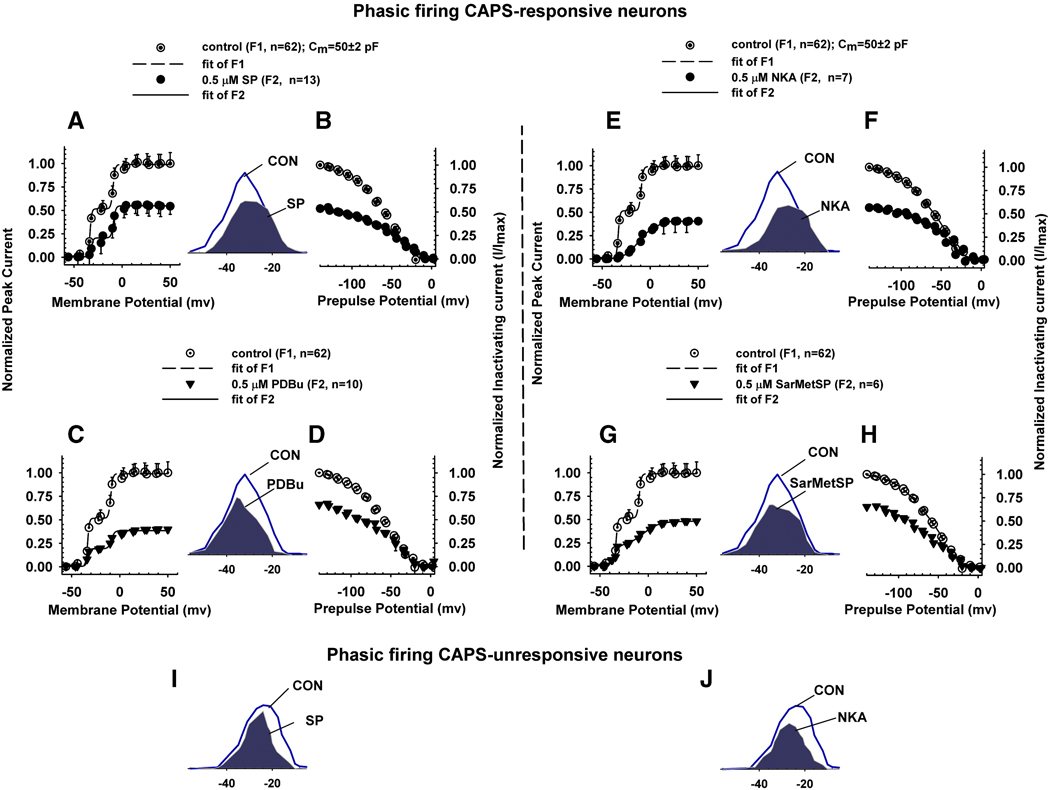
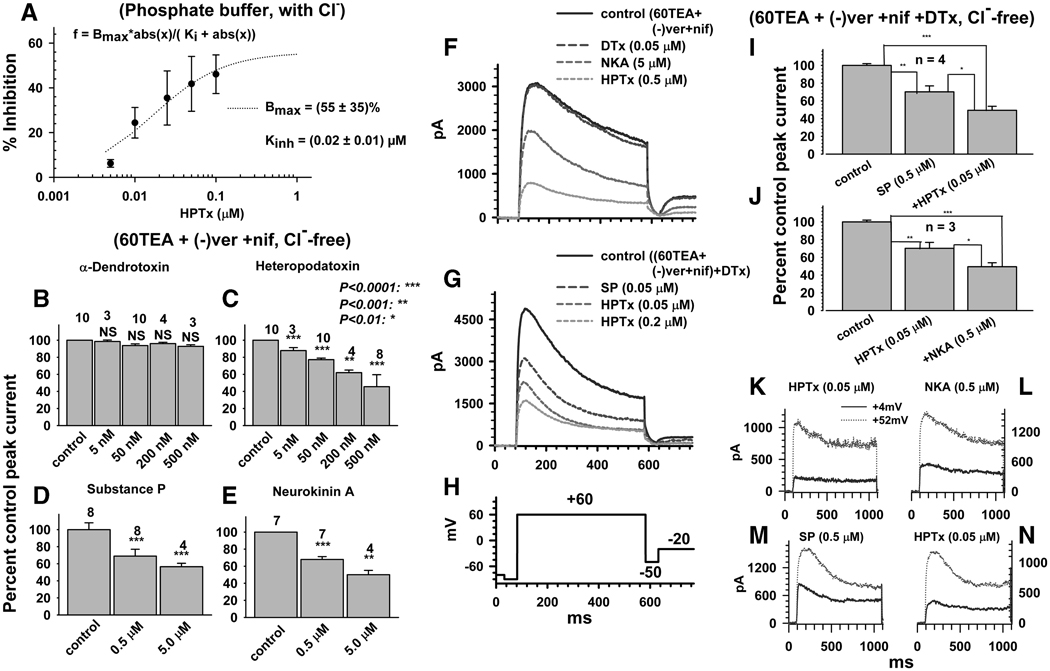
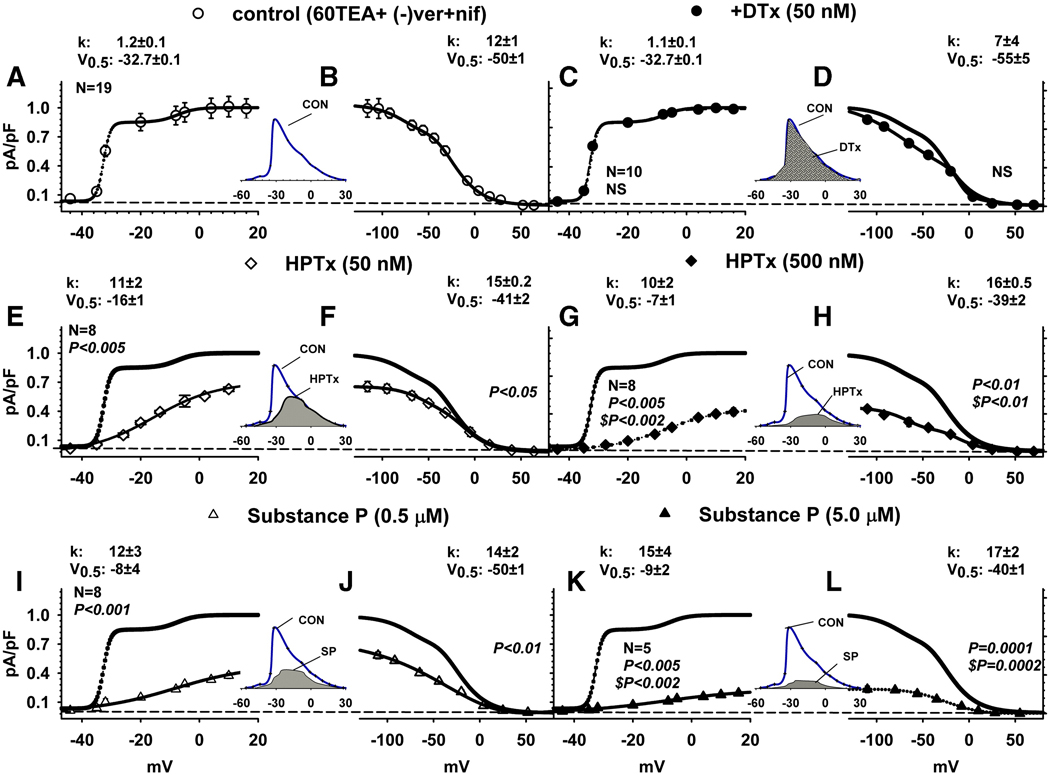
Neurokinins (NK) released from terminals of dorsal root ganglion (DRG) neurons may control firing of these neurons by an autofeedback mechanism. In this study we used patch clamp recording techniques to determine if NKs alter excitability of rat L4-S3 DRG neurons by modulating K(+) currents. In capsaicin (CAPS)-responsive phasic neurons substance P (SP) lowered action potential (AP) threshold and increased the number of APs elicited by depolarizing current pulses. SP and a selective NK(2) agonist, [betaAla(8)]-neurokinin A (4-10) also inhibited low threshold inactivating K(+) currents isolated by blocking non-inactivating currents with a combination of high TEA, (-) verapamil and nifedipine. Currents recorded under these conditions were heteropodatoxin-sensitive (Kv4 blocker) and alpha-dendrotoxin-insensitive (Kv1.1 and Kv1.2 blocker). SP and NKA elicited a >10 mV positive shift of the voltage dependence of activation of the low threshold currents. This effect was absent in CAPS-unresponsive neurons. The effect of SP or NKA on K(+) currents in CAPS-responsive phasic neurons was fully reversed by an NK(2) receptor antagonist (MEN10376) but only partially reversed by a PKC inhibitor (bisindolylmaleimide). An NK(1) selective agonist ([Sar(9), Met(11)]-substance P) or direct activation of PKC with phorbol 12,13-dibutyrate, did not change firing in CAPS-responsive neurons, but did inhibit various types of K(+) currents that activated over a wide range of voltages. These data suggest that the excitability of CAPS-responsive phasic afferent neurons is increased by activation of NK(2) receptors and that this is due in part to inhibition and a positive voltage shift in the activation of heteropodatoxin-sensitive Kv4 channels.
Figures
References
-
- Abdulla FA, Stebbing MJ, Smith PA. Effects of substance P on excitability and ionic currents of normal and axotomized rat dorsal root ganglion neurons. Eur. J. Neurosci. 2001;13:545–552. - PubMed
-
- Andersson KE. The overactive bladder: pharmacologic basis of drug treatment. Urol. 1997;50:74–84. - PubMed
-
- Andersson KE. Tachykinins: role in detrusor overactivity? Eur. Urol. 2006;49:423–425. - PubMed
-
- Averbeck B, Izydorczyk I, Kress M. Inflammatory mediators release calcitonin gene-related peptide from dorsal root ganglion neurons of the rat. Neurosci. 2000;98:135–140. - PubMed
-
- Besson JM. The neurobiology of pain. Lancet. 1999;353:1610–1615. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
