

doi: 10.1016/j.cell.2009.07.002.
Evolution of transcriptional regulatory circuits in bacteria
Affiliations
- PMID: 19632175
- PMCID: PMC2726713
- DOI: 10.1016/j.cell.2009.07.002
Item in Clipboard
Evolution of transcriptional regulatory circuits in bacteria
Cell.
.
Abstract
Related organisms typically respond to a given cue by altering the level or activity of orthologous transcription factors, which, paradoxically, often regulate expression of distinct gene sets. Although promoter rewiring of shared genes is primarily responsible for regulatory differences among related eukaryotic species, in bacteria, species-specific genes are often controlled by ancestral transcription factors, and regulatory circuit evolution has been further shaped by horizontal gene transfer. Modifications in transcription factors and in promoter structure also contribute to divergence in bacterial regulatory circuits.
Figures
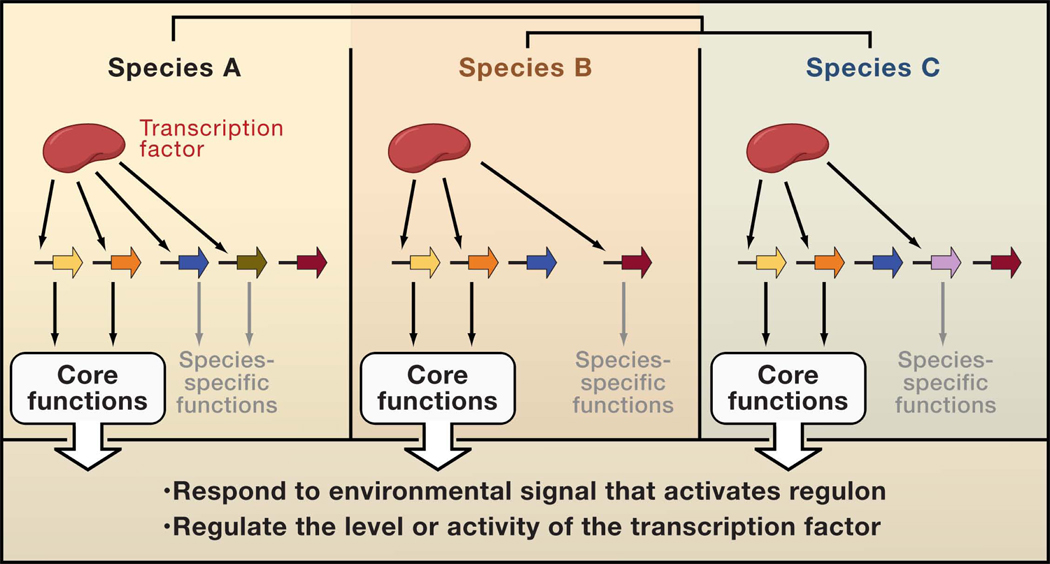
Depiction of three related bacterial species (termed A, B and C) sharing an ancestral transcription factor, which regulates variable gene sets in the three organisms. Only two target genes (yellow and orange arrows) are shared among the three species. The remaining target genes are regulated by the transcription factor in only one of the species because the target gene(s) is species-specific (green and lavender arrows) or because only one of the species harbors binding sites for the transcription factor in the promoter region of genes shared across species (brown and blue arrows.) The shared target genes, constituting the core regulon, have two main roles: to cope with the environmental change that activates the regulon, and to control the amount of the active form of the transcription factor. By contrast, the species-specific targets aid each species to proliferate in the particular niches in which they live.

Transcriptional regulatory circuits governing the expression of the pbgP gene mediate resistance to the antibiotic polymyxin B in enteric bacteria. The circuit in Y. pestis involves direct transcriptional control of the pbgP gene by the PhoP protein, which is activated when its cognate sensor PhoQ experiences low Mg2+, and by the PmrA protein, which is activated when its cognate sensor PmrB experiences Fe3+. The Y. pestis circuit, where the pbgP promoter harbors binding sites for the PhoP (blue box) and PmrA (red box) proteins, may constitute the ancestral state. The direct control of pbgP transcription by the PmrA protein in response to Fe3+ is shared by all depicted species, which differ in the activation of the pbgP gene taking place in low Mg2+. S. enterica utilizes an indirect pathway where the PhoP-dependent PmrD protein activates the PmrA protein post-translationally, enabling it to bind to the pbgP promoter and express pbgP in low Mg2+. There is no PhoP binding site in the S. enterica pbgP gene promoter. A PmrA binding site in the pmrD promoter mediates a negative feedback by the PmrA protein on the PhoP-promoted transcription of the pmrD gene. This feedback is believed to prevent the accumulation of the active (i.e., phosphorylated) form of the PmrA protein. K. pneumoniae’s circuit includes both direct pbgP control by the PhoP protein, like the Y. pestis’ circuit, as well as the indirect PmrD-mediated pathway operating in S. enterica. Yet, the PmrA protein does not exhibit negative feedback on pmrD transcription in K. pneumoniae. A hypothetical circuit lacking the latter negative feedback (shown to the left of the other circuits) is a hypothesized intermediate in the evolution from the circuit present in K. pneumoniae to the circuit operating in S. enterica. The pmrD gene was acquired (or invented) by the ancestral organism giving rise to K. pneumoniae, S. enterica and E. coli. Despite harboring a PhoP-activated pmrD gene, E. coli does not express PmrA-activated genes in low Mg2+ because it encodes a highly divergent PmrD protein. The E. coli PmrD protein may interact with and affect the activity of other regulatory protein(s). The most distant species depicted in this figure, E. coli and Y. pestis, diverged from their last common ancestor >200 million years ago (Deng et al., 2002).
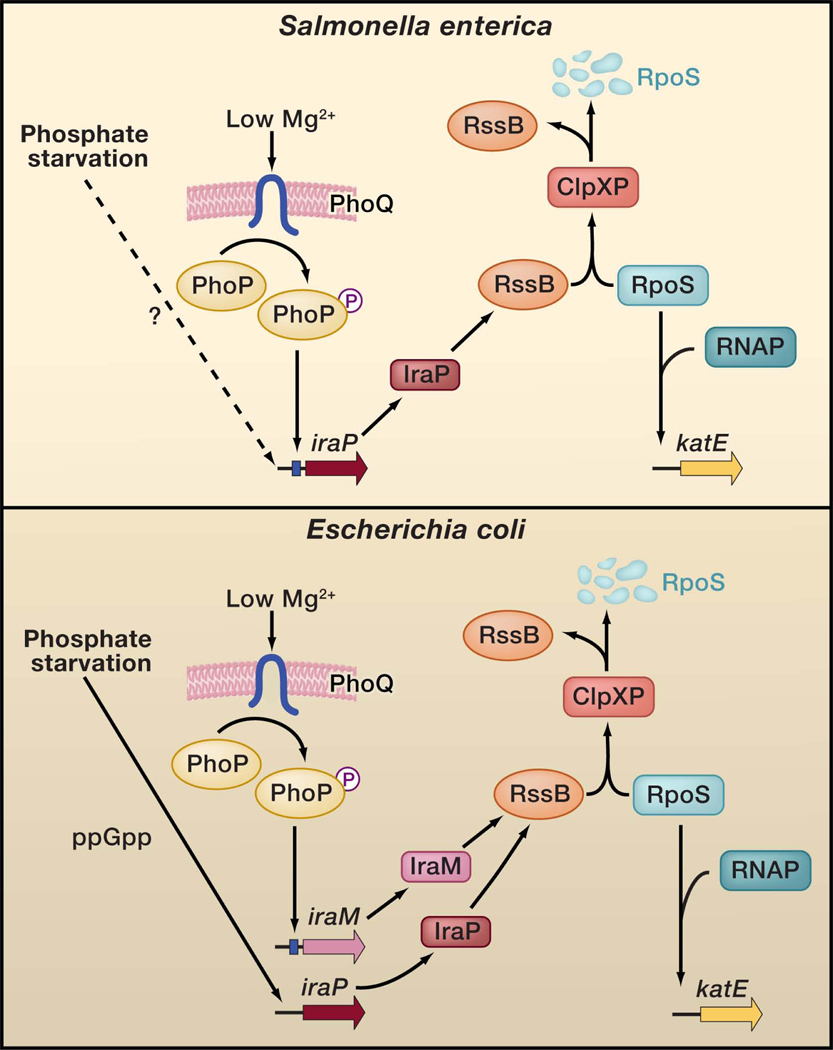
Promoter rewiring and a non-orthologous gene replacement distinguish the regulatory circuits controlling the stabilization of the alternative sigma factor RpoS taking place in low Mg2+ in Escherichia coli and Salmonella enterica. Under non-inducing conditions, the adaptor protein RssB (also designated SprE and MviA) delivers the RpoS protein to the ClpXP protease, which degrades RpoS. Specific nutrient-limiting conditions enhance RpoS protein levels. When E. coli or Salmonella experience phosphate limitation, the amount of the highly conserved IraP protein increase in a mechanism that is ppGpp-dependent in E. coli but uncharacterized in Salmonella. The IraP protein binds to the RssB protein, which is present in limiting amounts in the cell, thereby liberating RpoS to reprogram RNA polymerase (RNAP), which then transcribes RpoS-dependent promoters such as that corresponding to the katE gene. In low Mg2+ environments, IraP mediates RpoS accumulation in Salmonella but not in E. coli because the iraP promoter harbors a binding site (blue box) for the Mg2+-responsive PhoP protein in the former but not in the latter species. However, E. coli does accumulate RpoS and promote transcription of RpoS-regulated genes in the low Mg2+ conditions that activate the PhoP protein by utilizing the E. coli-specific PhoP-activated iraM gene. Despite exhibiting limited amino acid identity to one another, both IraP and IraM bind to and antagonize the RssB protein.

The structure of bacterial promoters dictates how transcription factors interact with RNA polymerase to promote transcription thereby impinging upon the evolution of regulatory circuits. A. The transcription factor binding sites participating in transcription initiation in bacteria are located typically < 100 nt upstream of the start site (+1). Within this region, they can lie either proximal or distal to the sites (small lavender boxes) recognized by RNA polymerase (composed ofβ, β’, 2α and σ subunits). Transcription factors (red) that bind in the proximal region (red bar) usually contact the σ or α N-terminal domain subunits of RNA polymerase. Transcription factors (blue) that bind in the distal region (blue bar) usually contact the flexible α C-terminal domain (CTD) subunit of RNA polymerase. Some transcription factors regulate promoters that harbor binding sites in either position, suggesting that they can make productive contacts with multiple RNA polymerase subunits. B. The transcription factor contact surface that is exposed to a given RNA polymerase subunit may vary depending on the orientation of a transcription factor binding site. This notion is consistent with reports for several transcription factors that their function is dependent on binding site orientation. New genes embedded in an ancestral regulon often harbor promoter structures distinct to those of ancestral members of the regulon (for example, the location and orientation of the transcription factor binding site are different even though the DNA motif is conserved). The location and orientation of a binding site can profoundly affect the ability of a transcription factor to regulate transcription. Thus, the regulation of distinct promoter structures probably demands the generation of new abilities in the transcription factor (such as gain of function mutations that allow it to interact with other RNA polymerase subunits).
References
-
- Alon U. Network motifs: theory and experimental approaches. Nat Rev Genet. 2007;8:450–461. - PubMed
-
- Balaji S, Aravind L. The two faces of short-range evolutionary dynamics of regulatory modes in bacterial transcriptional regulatory networks. Bioessays. 2007;29:625–629. - PubMed
-
- Beaber JW, Hochhut B, Waldor MK. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature. 2004;427:72–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
