

Mapping the cargo protein membrane translocation step into the PEX5 cycling pathway
- PMID: 19632994
- PMCID: PMC2785652
- DOI: 10.1074/jbc.M109.032565
Mapping the cargo protein membrane translocation step into the PEX5 cycling pathway
Abstract
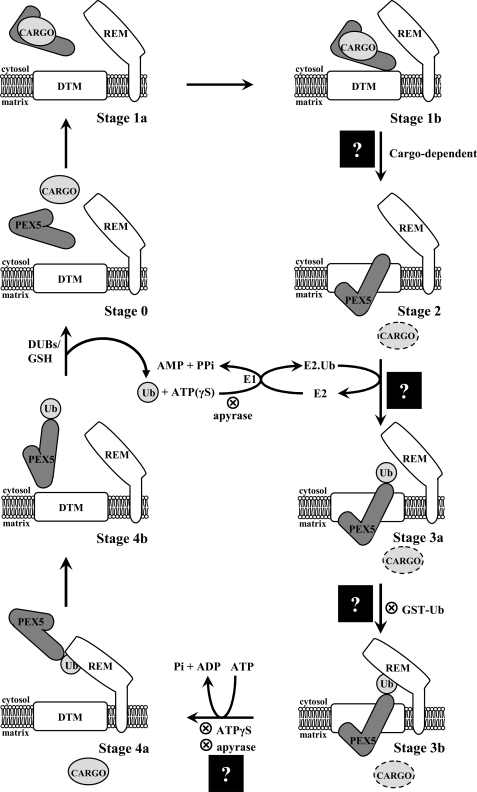
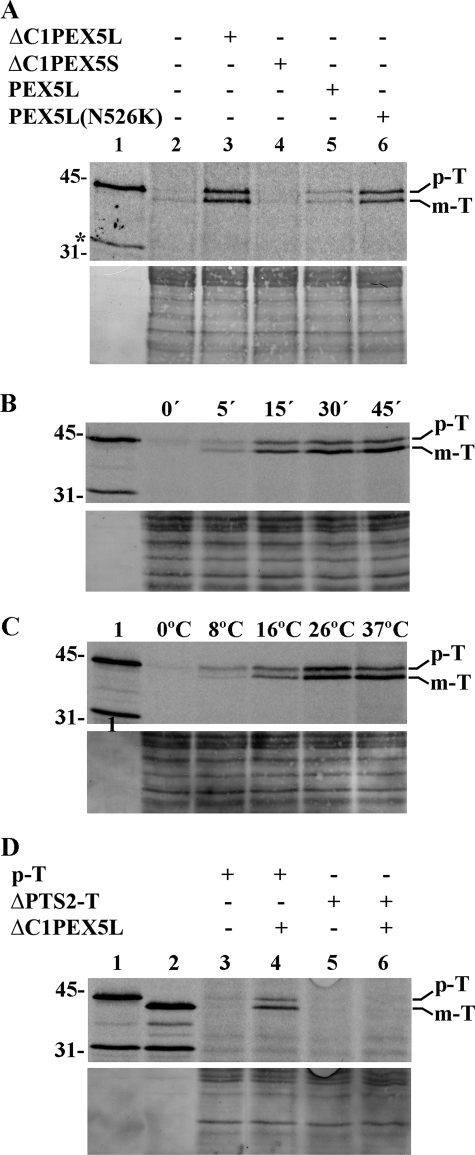
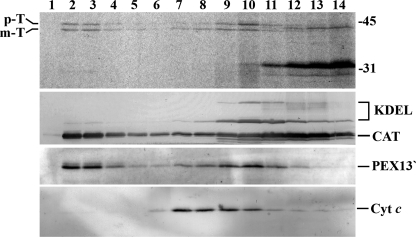
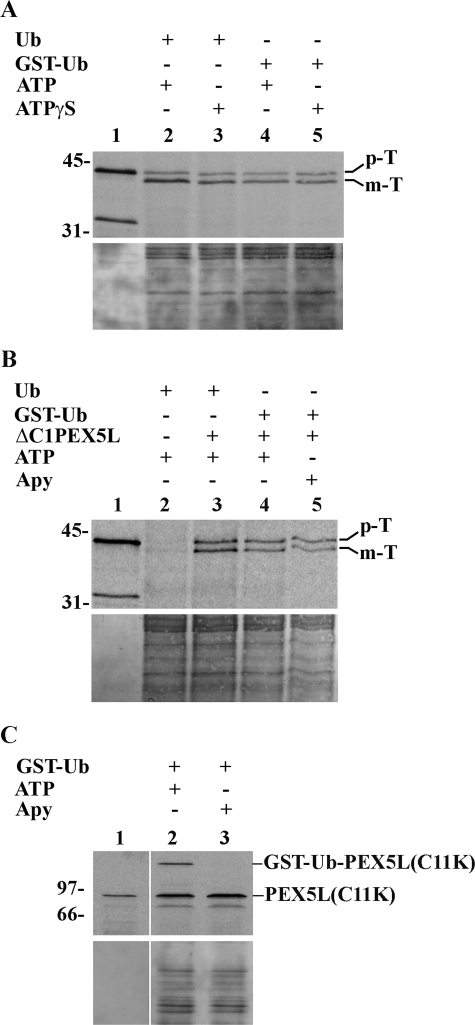
Newly synthesized peroxisomal matrix proteins are targeted to the organelle by PEX5, the peroxisomal cycling receptor. Over the last few years, valuable data on the mechanism of this process have been obtained using a PEX5-centered in vitro system. The data gathered until now suggest that cytosolic PEX5.cargo protein complexes dock at the peroxisomal docking/translocation machinery, where PEX5 becomes subsequently inserted in an ATP-independent manner. This PEX5 species is then monoubiquitinated at a conserved cysteine residue, a mandatory modification for the next step of the pathway, the ATP-dependent dislocation of the ubiquitin-PEX5 conjugate back into the cytosol. Finally, the ubiquitin moiety is removed, yielding free PEX5. Despite its usefulness, there are many unsolved mechanistic aspects that cannot be addressed with this in vitro system and that call for a cargo protein-centered perspective instead. Here we describe a robust peroxisomal in vitro import system that provides this perspective. The data obtained with it suggest that translocation of a cargo protein across the peroxisomal membrane, including its release into the organelle matrix, occurs prior to PEX5 ubiquitination.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
