

The apicobasal polarity kinase aPKC functions as a nuclear determinant and regulates cell proliferation and fate during Xenopus primary neurogenesis
- PMID: 19633170
- PMCID: PMC2730405
- DOI: 10.1242/dev.034454
The apicobasal polarity kinase aPKC functions as a nuclear determinant and regulates cell proliferation and fate during Xenopus primary neurogenesis
Abstract
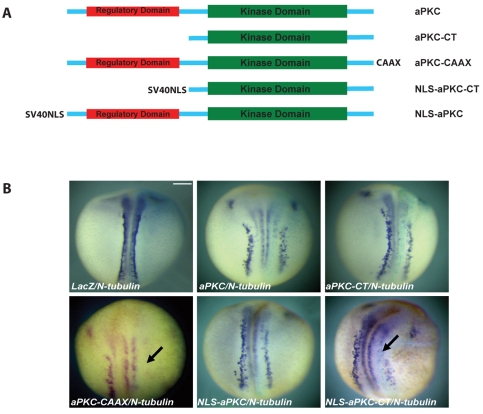
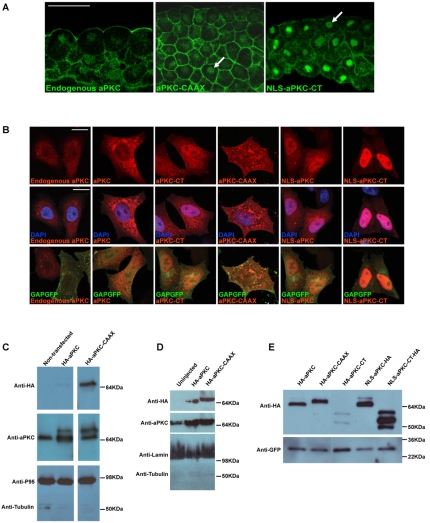
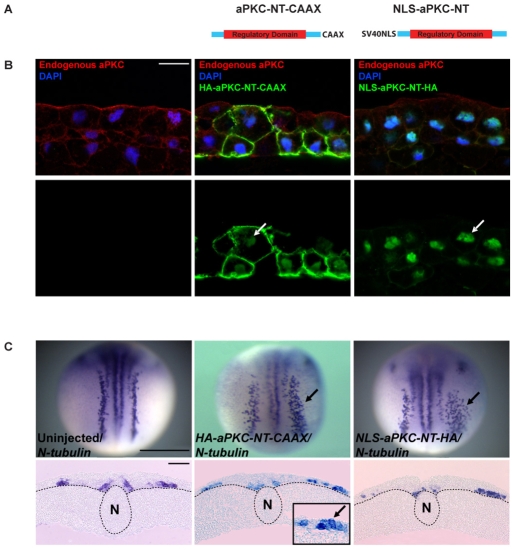
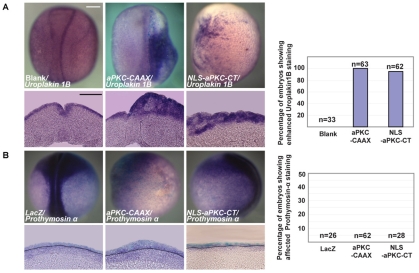
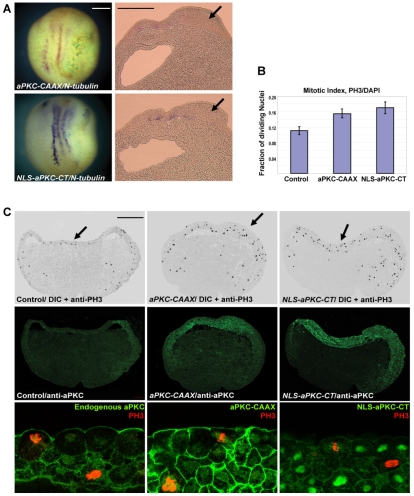
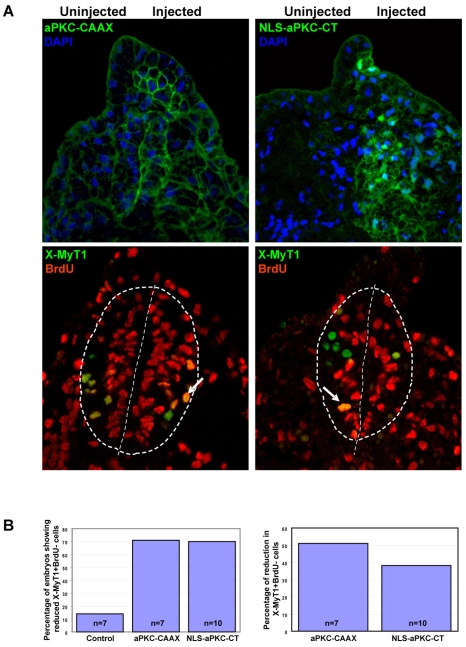
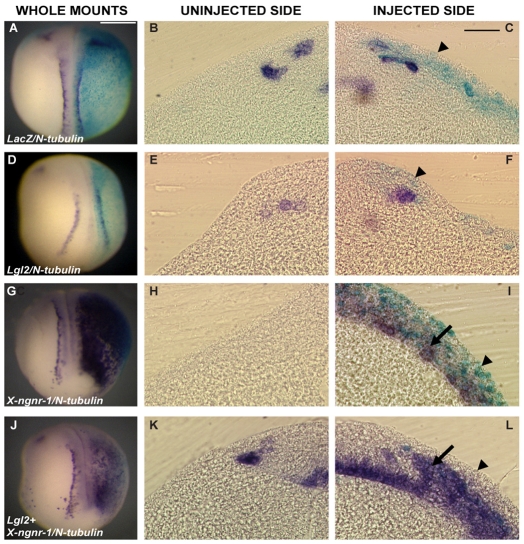
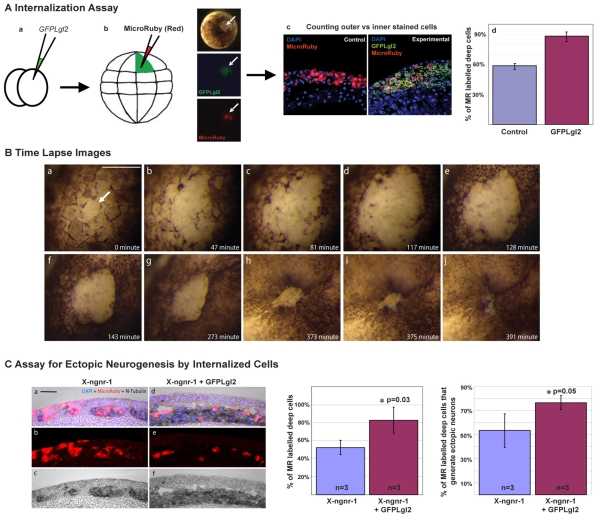
During neurogenesis in Xenopus, apicobasally polarised superficial and non-polar deep cells take up different fates: deep cells become primary neurons while superficial cells stay as progenitors. It is not known whether the proteins that affect cell polarity also affect cell fate and how membrane polarity information may be transmitted to the nucleus. Here, we examine the role of the polarity components, apically enriched aPKC and basolateral Lgl2, in primary neurogenesis. We report that a membrane-tethered form of aPKC (aPKC-CAAX) suppresses primary neurogenesis and promotes cell proliferation. Unexpectedly, both endogenous aPKC and aPKC-CAAX show some nuclear localisation. A constitutively active aPKC fused to a nuclear localisation signal has the same phenotypic effect as aPKC-CAAX in that it suppresses neurogenesis and enhances proliferation. Conversely, inhibiting endogenous aPKC with a dominant-negative form that is restricted to the nucleus enhances primary neurogenesis. These observations suggest that aPKC has a function in the nucleus that is important for cell fate specification during primary neurogenesis. In a complementary experiment, overexpressing basolateral Lgl2 causes depolarisation and internalisation of superficial cells, which form ectopic neurons when supplemented with a proneural factor. These findings suggest that both aPKC and Lgl2 affect cell fate, but that aPKC is a nuclear determinant itself that might shuttle from the membrane to the nucleus to control cell proliferation and fate; loss of epithelial cell polarity by Lgl2 overexpression changes the position of the cells and is permissive for a change in cell fate.
Figures
Similar articles
-
aPKC, Crumbs3 and Lgl2 control apicobasal polarity in early vertebrate development.Development. 2005 Mar;132(5):977-86. doi: 10.1242/dev.01645. Epub 2005 Feb 2. Development. 2005. PMID: 15689379
-
PAR1 specifies ciliated cells in vertebrate ectoderm downstream of aPKC.Development. 2007 Dec;134(23):4297-306. doi: 10.1242/dev.009282. Development. 2007. PMID: 17993468 Free PMC article.
-
Par6b regulates the dynamics of apicobasal polarity during development of the stratified Xenopus epidermis.PLoS One. 2013 Oct 18;8(10):e76854. doi: 10.1371/journal.pone.0076854. eCollection 2013. PLoS One. 2013. PMID: 24204686 Free PMC article.
-
aPKC phosphorylates p27Xic1, providing a mechanistic link between apicobasal polarity and cell-cycle control.Dev Cell. 2014 Dec 8;31(5):559-71. doi: 10.1016/j.devcel.2014.10.023. Dev Cell. 2014. PMID: 25490266 Free PMC article.
-
Lgl/aPKC and Crb regulate the Salvador/Warts/Hippo pathway.Fly (Austin). 2010 Oct-Dec;4(4):288-93. doi: 10.4161/fly.4.4.13116. Epub 2010 Oct 21. Fly (Austin). 2010. PMID: 20798605 Free PMC article. Review.
Cited by
-
aPKC in neuronal differentiation, maturation and function.Neuronal Signal. 2019 Sep;3(3):NS20190019. doi: 10.1042/NS20190019. Epub 2019 Sep 23. Neuronal Signal. 2019. PMID: 32269838 Free PMC article. Review.
-
Fezf2 promotes neuronal differentiation through localised activation of Wnt/β-catenin signalling during forebrain development.Development. 2014 Dec;141(24):4794-805. doi: 10.1242/dev.115691. Development. 2014. PMID: 25468942 Free PMC article.
-
Atypical protein kinase C couples cell sorting with primitive endoderm maturation in the mouse blastocyst.Development. 2013 Nov;140(21):4311-22. doi: 10.1242/dev.093922. Epub 2013 Sep 25. Development. 2013. PMID: 24067354 Free PMC article.
-
Analysis of neural progenitors from embryogenesis to juvenile adult in Xenopus laevis reveals biphasic neurogenesis and continuous lengthening of the cell cycle.Biol Open. 2015 Nov 30;4(12):1772-81. doi: 10.1242/bio.013391. Biol Open. 2015. PMID: 26621828 Free PMC article.
-
Rare variants in PRKCI cause Van der Woude syndrome and other features of peridermopathy.medRxiv [Preprint]. 2025 Jan 17:2025.01.17.25320742. doi: 10.1101/2025.01.17.25320742. medRxiv. 2025. Update in: Am J Hum Genet. 2025 Aug 29:S0002-9297(25)00318-0. doi: 10.1016/j.ajhg.2025.08.008. PMID: 39867391 Free PMC article. Updated. Preprint.
References
-
- Asashima, M. and Grunz, H. (1983). Effects of inducers on inner and outer gastrula ectoderm layers of Xenopus laevis. Differentiation 23, 206-212. - PubMed
-
- Bellefroid, E. J., Bourguignon, C., Hollemann, T., Ma, Q., Anderson, D. J., Kintner, C. and Pieler, T. (1996). X-MyT1, a Xenopus C2HC-type zinc finger protein with a regulatory function in neuronal differentiation. Cell 87, 1191-1202. - PubMed
-
- Betschinger, J. and Knoblich, J. A. (2004). Dare to be different: asymmetric cell division in Drosophila, C. elegans and vertebrates. Curr. Biol. 14, R674-R685. - PubMed
-
- Bourguignon, C., Li, J. and Papalopulu, N. (1998). XBF-1, a winged helix transcription factor with dual activity, has a role in positioning neurogenesis in Xenopus competent ectoderm. Development 125, 4889-4900. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
