

Basal lamina strengthens cell membrane integrity via the laminin G domain-binding motif of alpha-dystroglycan
- PMID: 19633189
- PMCID: PMC2715328
- DOI: 10.1073/pnas.0906545106
Basal lamina strengthens cell membrane integrity via the laminin G domain-binding motif of alpha-dystroglycan
Abstract
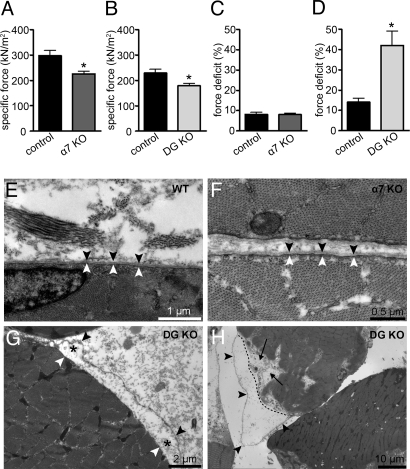
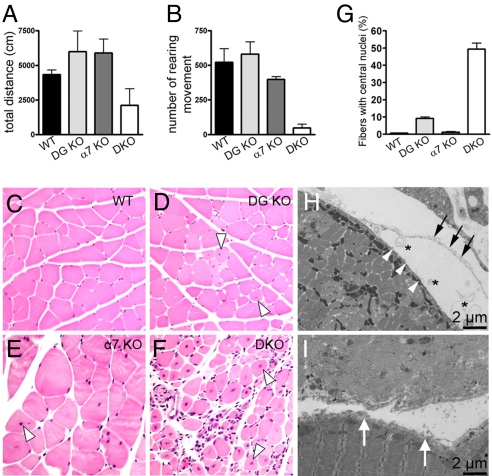
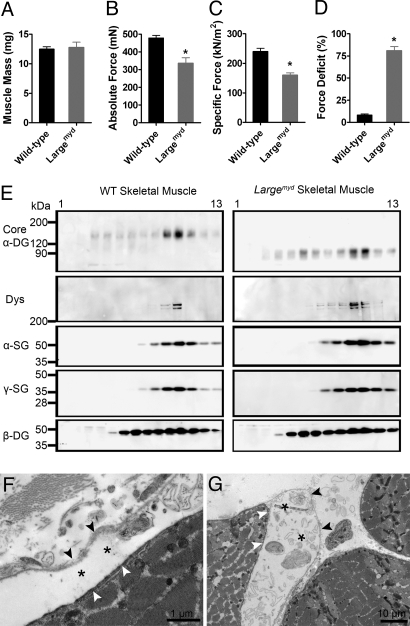
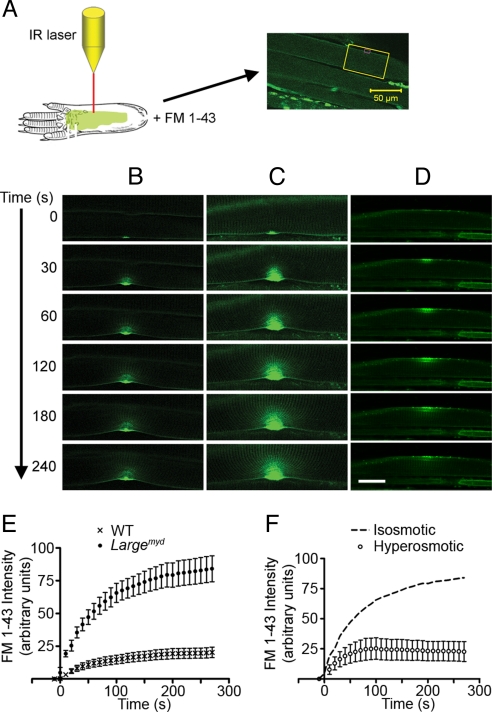
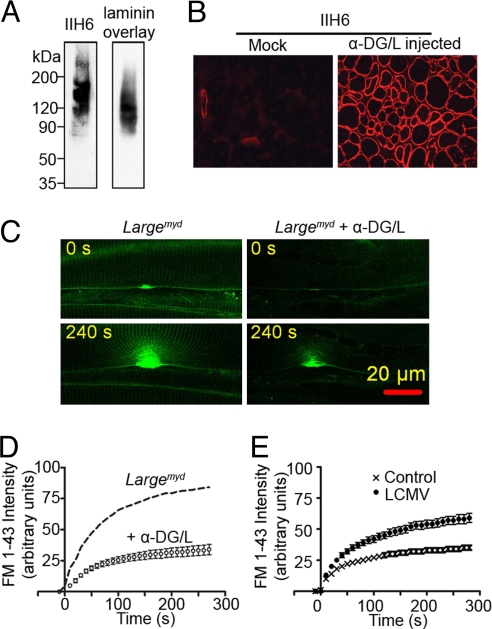
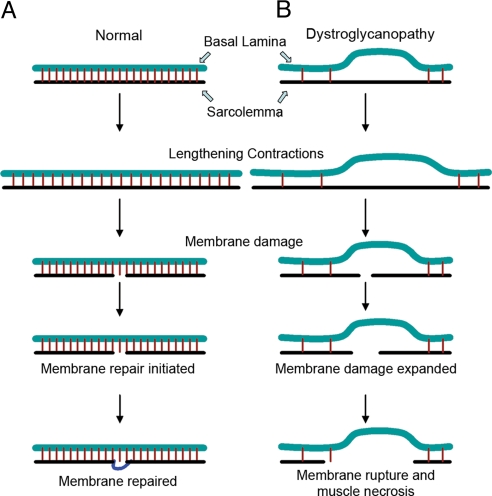
Skeletal muscle basal lamina is linked to the sarcolemma through transmembrane receptors, including integrins and dystroglycan. The function of dystroglycan relies critically on posttranslational glycosylation, a common target shared by a genetically heterogeneous group of muscular dystrophies characterized by alpha-dystroglycan hypoglycosylation. Here we show that both dystroglycan and integrin alpha7 contribute to force-production of muscles, but that only disruption of dystroglycan causes detachment of the basal lamina from the sarcolemma and renders muscle prone to contraction-induced injury. These phenotypes of dystroglycan-null muscles are recapitulated by Large(myd) muscles, which have an intact dystrophin-glycoprotein complex and lack only the laminin globular domain-binding motif on alpha-dystroglycan. Compromised sarcolemmal integrity is directly shown in Large(myd) muscles and similarly in normal muscles when arenaviruses compete with matrix proteins for binding alpha-dystroglycan. These data provide direct mechanistic insight into how the dystroglycan-linked basal lamina contributes to the maintenance of sarcolemmal integrity and protects muscles from damage.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Davies KE, Nowak KJ. Molecular mechanisms of muscular dystrophies: Old and new players. Nat Rev Mol Cell Biol. 2006;7:762–773. - PubMed
-
- Durbeej M, Campbell KP. Muscular dystrophies involving the dystrophin-glycoprotein complex: An overview of current mouse models. Curr Opin Genet Dev. 2002;12:349–361. - PubMed
-
- Sanes JR. The basement membrane/basal lamina of skeletal muscle. J Biol Chem. 2003;278:12601–12604. - PubMed
-
- Candiello J, et al. Biomechanical properties of native basement membranes. FEBS J. 2007;274:2897–2908. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
