

The tRNA modification complex elongator regulates the Cdc42-dependent mitogen-activated protein kinase pathway that controls filamentous growth in yeast
- PMID: 19633267
- PMCID: PMC2747823
- DOI: 10.1128/EC.00015-09
The tRNA modification complex elongator regulates the Cdc42-dependent mitogen-activated protein kinase pathway that controls filamentous growth in yeast
Abstract
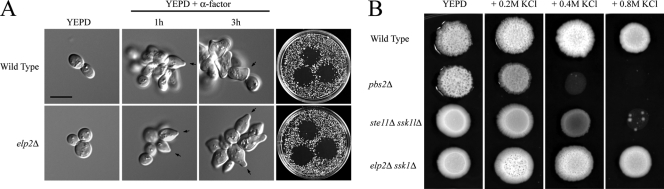
Signal transduction pathways control multiple aspects of cellular behavior, including global changes to the cell cycle, cell polarity, and gene expression, which can result in the formation of a new cell type. In the budding yeast Saccharomyces cerevisiae, the mitogen-activated protein kinase (MAPK) pathway that controls filamentous growth induces a dimorphic foraging response under nutrient-limiting conditions. How nutritional cues feed into MAPK activation remains an open question. Here we report a functional connection between the elongator tRNA modification complex (ELP genes) and activity of the filamentous growth pathway. Elongator was required for filamentous growth pathway signaling, and elp mutants were defective for invasive growth, cell polarization, and MAPK-dependent mat formation. Genetic suppression analysis showed that elongator functions at the level of Msb2p, the signaling mucin that operates at the head of the pathway, which led to the finding that elongator regulates the starvation-dependent expression of the MSB2 gene. The Elp complex was not required for activation of related pathways (pheromone response or high osmolarity glycerol response) that share components with the filamentous growth pathway. Because protein translation provides a rough metric of cellular nutritional status, elongator may convey nutritional information to the filamentous growth pathway at the level of MSB2 expression.
Figures





Similar articles
-
Cdc42p-interacting protein Bem4p regulates the filamentous-growth mitogen-activated protein kinase pathway.Mol Cell Biol. 2015 Jan;35(2):417-36. doi: 10.1128/MCB.00850-14. Epub 2014 Nov 10. Mol Cell Biol. 2015. PMID: 25384973 Free PMC article.
-
Functions for Cdc42p BEM adaptors in regulating a differentiation-type MAP kinase pathway.Mol Biol Cell. 2020 Mar 15;31(6):491-510. doi: 10.1091/mbc.E19-08-0441. Epub 2020 Jan 15. Mol Biol Cell. 2020. PMID: 31940256 Free PMC article.
-
Possible integration of upstream signals at Cdc42 in filamentous differentiation of S. cerevisiae.Yeast. 2005 Oct 15;22(13):1069-77. doi: 10.1002/yea.1294. Yeast. 2005. PMID: 16200521
-
Structures and Activities of the Elongator Complex and Its Cofactors.Enzymes. 2017;41:117-149. doi: 10.1016/bs.enz.2017.03.001. Epub 2017 Apr 12. Enzymes. 2017. PMID: 28601220 Review.
-
SSD1 modifies phenotypes of Elongator mutants.Curr Genet. 2020 Jun;66(3):481-485. doi: 10.1007/s00294-019-01048-9. Epub 2019 Nov 27. Curr Genet. 2020. PMID: 31776648 Free PMC article. Review.
Cited by
-
Unfolded Protein Response Suppression in Yeast by Loss of tRNA Modifications.Genes (Basel). 2018 Oct 23;9(11):516. doi: 10.3390/genes9110516. Genes (Basel). 2018. PMID: 30360492 Free PMC article.
-
Multiple signals converge on a differentiation MAPK pathway.PLoS Genet. 2010 Mar 19;6(3):e1000883. doi: 10.1371/journal.pgen.1000883. PLoS Genet. 2010. PMID: 20333241 Free PMC article.
-
Gene by Environment Interactions reveal new regulatory aspects of signaling network plasticity.PLoS Genet. 2022 Jan 4;18(1):e1009988. doi: 10.1371/journal.pgen.1009988. eCollection 2022 Jan. PLoS Genet. 2022. PMID: 34982769 Free PMC article.
-
Comparative Transcriptome Analysis Identified Candidate Genes Related to Bailinggu Mushroom Formation and Genetic Markers for Genetic Analyses and Breeding.Sci Rep. 2017 Aug 24;7(1):9266. doi: 10.1038/s41598-017-08049-z. Sci Rep. 2017. PMID: 28839254 Free PMC article.
-
The acyl-CoA-binding protein VdAcb1 is essential for carbon starvation response and contributes to virulence in Verticillium dahliae.aBIOTECH. 2024 Jul 13;5(4):431-448. doi: 10.1007/s42994-024-00175-3. eCollection 2024 Dec. aBIOTECH. 2024. PMID: 39650135 Free PMC article.
References
-
- Bouwmeester, T., A. Bauch, H. Ruffner, P. O. Angrand, G. Bergamini, K. Croughton, C. Cruciat, D. Eberhard, J. Gagneur, S. Ghidelli, C. Hopf, B. Huhse, R. Mangano, A. M. Michon, M. Schirle, J. Schlegl, M. Schwab, M. A. Stein, A. Bauer, G. Casari, G. Drewes, A. C. Gavin, D. B. Jackson, G. Joberty, G. Neubauer, J. Rick, B. Kuster, and G. Superti-Furga. 2004. A physical and functional map of the human TNF-α/NF-κB signal transduction pathway. Nat. Cell Biol. 6:97-105. - PubMed
-
- Carbone, A., A. Zinovyev, and F. Kepes. 2003. Codon adaptation index as a measure of dominating codon bias. Bioinformatics 19:2005-2015. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
